| |
16) Physiological data
Whatever relevant physiologic data are available have been provided in review
form by Silberman & Fulton (1979), and as bibliography by Miller (1983).
17)
Other resources
Cell strains of four species of rhinoceros are available from the Frozen zoo at the Zoological Society of San Diego by contacting Dr. Oliver Ryder at oryder@ucsd.edu.
None are available of the severely endangered Javan rhinoceros.
References
Ashley,
M.V., Melnick, D.J. and Western, D.: Conservation genetics of the black
rhinoceros (Diceros bicornis): I. Evidence from the mitochondrial DNA
of 3 populations. Conserv. Biol. 1:71-77, 1990.
Benirschke,
K. and Lowenstine, L.J.: The placenta of the rhinocerotidae. Verh. Ber.
Erkr. Zootiere (Dresden). 37:15-23, 1995.
Carlstead,
K., Fraser, J., Bennett, C. and Kleiman, D.G.: Black rhinoceros (Diceros
bicornis) in U.S. Zoos: II. Behavior, breeding success, and mortality
in relation to housing facilities. Zoo Biol. 18:35-52, 1999.
Chapin,
H., Malecek, A.C., Miller, R.E., Bell, C.E., Gray, L.S. and Hunter, V.L.:
Acute intravascular hemolytic anemia in the black rhinoceros: Hematologic
and immunohematologic observations. Amer. J. Vet. Med. 47:1313-1320, 1986.
Dolinar,
Z.J., Ludwig, K.S. und Müller, E.: Ein weiterer Beitrag zur Kenntnis
der Placenten der Ordnung Perissodactyla: Zwei Geburtsplacenten des Indischen
Panzernashorns. (Rhinoceros unicornis L.). Acta Anat. 61:331-354, 1965.
Galama, W.T., Graham, L.H. and Savage, A.: Comparison of fecal storage methods for steroid analysis in black rhinoceros (Diceros bicornis). Zoo Biol. 23:291-300, 2004.
George,
M. Jr., Chemnick, L.G., Cisova, D., Gabrisova, E., Stratil, A. and Ryder,
O.A.: Genetic differentiation of white rhinoceros subspecies: diagnostic
differences in mitochondrial DNA and serum proteins. In, Proc. Intern.
Conference on Rhinoceros Biology and Conservation, San Diego, CA 1991,
pp. 105-113.
Groves,
C.P.: Taxonomic notes on the white rhinoceros. Ceratotherium simum (Burchell,
1817). Säugetierk. Mitteil. 23:200-212, 1975.
Groves,
C.P.: Phylogeny of the living species of Rhinoceros. Z. zool. System.
Evol. 21:293-313, 1983.
Hansen, K.M.: Q-bands of some chromosomes of white rhinoceros (Diceros
simus). Hereditas 82:205-208, 1976.
Harley,
E.H. and O'Ryan, C.: Molecular genetic studies of southern African rhinoceros.
In, Proc. Intern. Conference on Rhinoceros Biology and Conservation, San
Diego, CA 1991, pp. 101-104.
Heinichen,
I.G.: Karyological studies on southern African perissodactyla. Kodoe 13:51-108,
1970.
Houck,
M.L., Ryder, O.A., Váhala, J., Kock, R.A. and Oosterhuis, J.E.:
Diploid chromosome number and chromosomal variation in the white rhinoceros
(Ceratotherium simum). J. Hered. 85:30-34, 1994.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 7:Folio 339, 1973. Springer-Verlag, New York.
Hungerford,
D.A., Chandra, H.S. and Snyder, R.L.: Somatic chromosomes of a black rhinoceros
(Diceros bicornis Gray 1821). Amer. Naturalist 101:357-358, 1967.
Lang,
E.M.: Geburt eines Panzernashorns, Rhinoceros unicornis, im Zoologischen
Garten Basel. Säugetierk. Mitt. 5:69-70, 1957.
Lang,
E.M.: Einige biologische Daten vom Panzernashorn (Rhinoceros unicornis).
Rev. Suisse Zool. Genève 74:603-607, 1967.
Laurie,
W,A., Lang, E.M. and Groves, C.P.: Rhinoceros unicornis. In, Mammalian
Species # 211, pp.1-6, 1983. Amer. Soc. Mammalogy.
Ludwig,
K.S.: Zur Kenntnis der Geburtsplacenten der Ordnung Perissodactyla. Acta
Anat. 49:154-167, 1962.
Ludwig,
K.S. und Villiger, W.: Zur Ultrastruktur der Blattzottenepithelien in der
Placenta des Indischen Panzernashorns (Rhinoceros unicornis L.). Acta Anat.
62:593-605, 1965.
Ludwig, K.S. und Müller, E.: Zur Histochemie der Placenta des Panzernashorns
(Rhinoceros unicornis L.). Acta Anat. Suppl. 115:155-159, 1965.
Mereniender,
A.M., Woodruff, D.S., Ryder, O.A., Kock, R. and Váhala, J.: Allozyme
variation and differentiation in African and Indian rhinoceroses. J. Hered.
80:377-382, 1989.
Miller, R.E.: Veterinary Bibliography for Rhinoceros. A.A. Balkema Publ.,
Amsterdam , 1983.
Miller,
R.E. and Boever, W.J.: Fatal hemolytic anemia in the black rhinoceros:
Case report and a survey. J.A.V.M.A. 181:1228-1231, 1982.
Morales,
J.C., Andau, P.M., Supriatna, J., Zainuddi, Z.Z. and Melnick, D.J.: Mitochondrial
DNA variability and conservation genetics of the Sumatran rhinoceros.
Conserv. Biol. 11:539-543, 1997.
Paglin,
D.E., Valentine, W.N., Miller, R.E., Nakatani, M. and Brockway, R.A.:
Acute intravascular hemolysis in the black rhinoceros: Erythrocyte enzymes
and metabolic intermediates. Amer. J. Vet. Res. 47:1321-1325, 1986.
Patton,
L., Czekala, N. and Lance, V.: Workshop on problems associated with the
low rate of reproduction among captive-born female Southern White Rhinoceros
(Ceratotherium simum simum). Zoological Society of San Diego, 1998.
Patton, M.L., Swaisgood, R.R., Czekala , N.M. , White, A.M., Fetter, G.A., Montagne, J.P., Rieches, R.G. and Lance, V.A.: Reproductive cycle length and pregnancy in the Southern white rhinoceros (Ceratotherium simum simum) as determined by fecal pregnane analysis and observations of mating behavior. Zoo Biol. 18:111-127, 1999.
Radcliffe,
R.W., Czekala, N.M. and Osofsky, S.A.: Technical Report. Combined serial
ultrasonography and fecal progestin analysis for reproductive evaluation
of the female white rhinoceros (Ceratotherium simum simum): Preliminary
results. Zoo Biol. 16:445-456, 1997.
Ramsey, E.: The Placenta of Laboratory Animals and Man. Holt, Rinehart
and Winston, NY 1975.
Ryder,
O.A., ed.: Rhinoceros Biology and Conservation. Zoological Society of San
Diego, 1993 (Proc. of Conference 1991).
Schaller,
K. and Pilaski, J.: Pocken bei Breitmaul-nashörnern (Ceratotherium
s. simum) im Zoologischen Garten Münster. Zool. Garten 49:169-184,
1979.
Silberman,
M.S. and Fulton, R.B.: Medical problems of captive and wild rhinoceros
- a review of the literature and personal experiences. J. Zoo Anim. Med.
10:6-16, 1979.
Wurster, D.H. and Benirschke, K.: The chromosomes of the Great Indian
Rhinoceros (Rhinoceros unicornis L.). Experientia 24:511, 1968.
Groves, C.P.: Ceratotherium simum. In, Mammalian Species. 8:1-6, 1972.
Amer. Soc. Mammalogy.
.
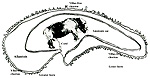
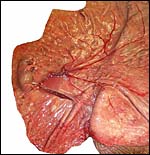
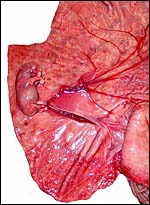
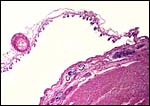
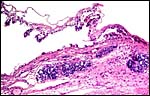
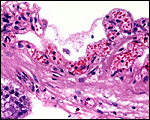
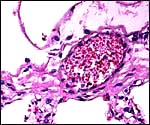
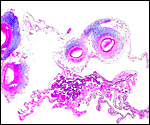
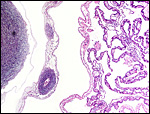
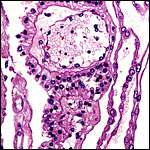
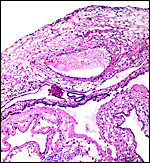
18) Other data to be accumulated
There is a need for the delineation of hormonal profiles during pregnancy
because of the fetal gonadal stimulation. The length of umbilical cords
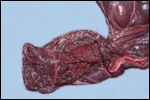
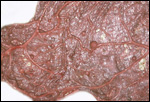
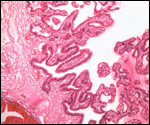
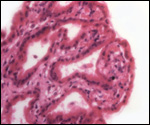
is not securely established. No implanted placenta has been observed and
thus no definitive statements concerning the fetal/maternal interface
can be made.
|