| |
13)
Genetics
Thomson's gazelles have 58 chromosomes in both sexes (Nelson-Rees et al.,
1967; Wurster & Benirschke, 1968). They lack the X/autosome translocation
found in so many other African antilopinae (Effron et al., 1976).
A male hybrid with Gazella rufifrons was born in London but died
the next day (Gray, 1972)
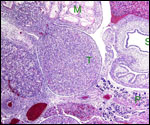
14) Immunology
I know of no immunological studies.
15)
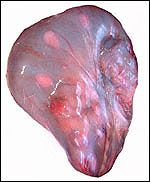
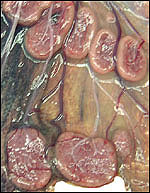
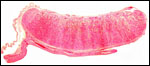
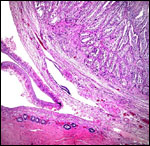
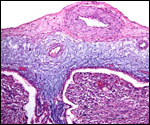
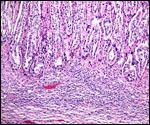
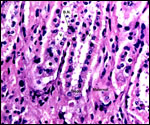
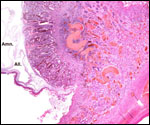
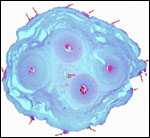
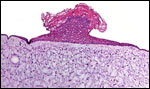
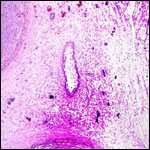
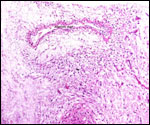
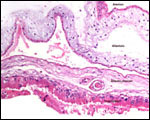
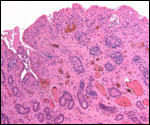
Pathological features
Trauma was found to be the commonest cause of mortality (Griner, 1983).
16)
Physiologic data
No data are known to me.
17)
Other resources
Cell strains of this gazelle species are available from CRES
at San Diego zoo by contacting Dr. Oliver Ryder at: oryder@ucsd.edu.
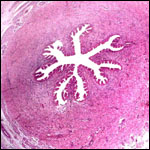
18) Other remarks - What additional Information is needed?
No weights of mature placentas, nor have the lengths of their term gestation
umbilical cords been recorded. Endocrine studies of gestation are needed.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society
of San Diego. I appreciate also very much the help of the pathologists
at the San Diego Zoo.
References
Effron, M., Bogart, M.H., Kumamoto, A.T. and Benirschke, K.: Chromosome
studies in the mammalian subfamily Antilopinae. Genetica 46:419-444,
1976.
Estes,
R.D.: The comparative behavior of Grant's and Thomson's gazelles. J. Mammal.
48:189-209, 1967.
Gray,
A.P.: Mammalian Hybrids. A Check-list with Bibliography. 2nd edition.
Commonwealth Agricultural Bureaux Farnham Royal, Slough, England, 1972.
Griner,
L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego,
California, 1983.
Jones,
M.L.: Longevity of ungulates in captivity. Int. Zoo Ybk. 32:159-169, 1993.
Kellas,
L.M.: An intra-epithelial granular cell in the uterine epithelium of some
ruminant species during the pregnancy cycle. Acta Anat. 44:109-xxx, 1961.
Kellas,
L.M.: The placenta and foetal membranes of the antelope Ourebia ourebi
(Zimmermann). Acta Anat. 64:390-445, 1966.
Krolling,
O.: Über den Bau der Antilopenplazentome. Z. Mikrosk. Anat. Forsch.
27:216-232, 1931.
Matthee,
C.R. and Robinson, T.J.: Cytochrome b phylogeny of the family bovidae:
Resolution within the alcelaphini, antilopini, neotragini, and tragelaphini.
Molec. Phylogenet. Evol. 12:31-46, 1999.
Mentis,
M.T.: A review of some life history features of the large herbivores of
Africa. The Lammergeyer 1-89, 1972.
Nelson-Rees,
W.A., Kniazeff, A.J., Darby, N.B. and Malley, R.I.: Chromosomes of a male
gazelle. Gazella thomsoni and a female tapir. Tapirus terrestris
columbianus. Mamm. Chromosomes Newsl. 8:229-230, 1967.
Nowak,
R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press,
Baltimore, 1999.
Walther,
F.R.: Territorial behaviour in certain horned ungulates, with special
reference to the examples of Thomson's and Grant's gazelles. Zoologica
Africana 7:303-307, 1972.
Wurster,
D.H. and Benirschke, K.: Chromosome studies in the superfamily Bovoidea.
Chromosoma 25:152-171, 1968.
|