 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last
updated: |
Raphicerus campestris
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
This genus contains three species (R. campestris, R. melanotis, R. sharpei), all small, shy antelopes of Africa with several subspecies. Raphis = needle; ceros (keras) = horn (Gotch, 1979). They are widely distributed in Southern Africa and, being browsers, they are said to be in need of some cover but are otherwise in need of only very little water (Nowak, 1999). Only the males have short horns and all have conspicuous pre-orbital glands. Their lifespan in captivity is 8+ years according to Jones (1993) and they weigh between 6 and 16 kg as adults. [steen = stone - this is perhaps a "perversion" of the original settlers who named animals according to what they knew from the past, in this case "Steinbock", but "brick buck" in Afrikaans refers to the reddish color of the animal. The animals do not prefer rocks but flat savannah]. A common belief is that, when feeling endangered, steenbok seeks refuge in the holes of aardvarks. Vrba & Schaller (2000) indicated that the first appearance of Raphicerus dates back 4.5-5 MYA.
 |
Young male steenbok at San Diego Zoo, newly arrived. |
 |
Female steenbok at San Diego Zoo. |
Single young weighing around 1,000 g are born after a gestation that lasts around 7 months (168-210 days! Nowak, 1999). Mentis (1972) and others mention sources that suggest occasional twins to be born. The most comprehensive reproductive study undertaken is probably that by Dasmann & Mossman (1962).
3) Implantation
No early implantations have been described.
Steenbok have a multicotyledonary placenta with four distinct rows of cotyledons that correspond to the caruncles. The feto-maternal relationship is typically epitheliochorial. There is no invasion of the endometrium by trophoblast. In addition to the descriptions by Hradecky et al. (1983; 1988), I have had one placenta available to me. The right uterine horn contained 18 cotyledons, the left horn had 26 (= 44 cotyledons total). The individual cotyledons measured up to 3 x 2 cm in diameters. The specimen comes from a dead pregnant animal that contained a fetus with 15.5 cm CR length. It is interesting that Hradecky (1983) found 71 cotyledons in one and 63 (between 17 and 92) cotyledons in a very large number of cases (# 131). He did not describe their location.
In all respects, this placenta resembles closely the placenta of the Oribi. That species' placenta has been described in exquisite detail and depicted by Kellas (1966).
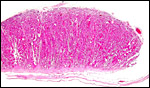 |
Section of implanted steenbok cotyledon with thin uterus below. Note the endometrial extensions in between the villi. |
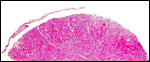 |
Horizontal section of steenbok cotyledon, taken through the villous tissue. |
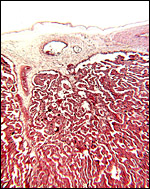 |
Fetal surface with chorion and villi branching between villi. |
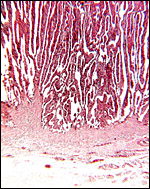 |
Floor of implanted cotyledon; the myometrium in below and the extensions of the fibrous endometrium extend up between the slender villi. |
This is a typical epithelio-chorial placenta. The trophoblast is cuboidal and has numerous typical binucleate cells. No true syncytium is evident and there is a minimal amount of black pigment in areas of subchorionic trophoblast but it may be artifact. At least none of the brown pigment found in other artiodactyla is present.
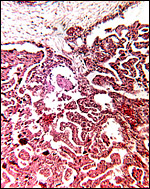 |
Chorion is above and sparsely-branched slender villi interdigitate with endometrium. |
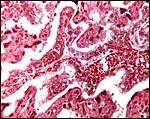 |
Low power view of villous tissue interdigitating with endometrium. |
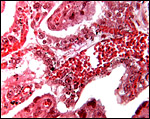 |
Large vessel within villus whose trophoblast is adjacent to endometrial invagination. Occasional binucleate cells are present. |
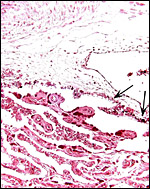 |
The minimal amount of black pigment in subchorionic trophoblast is arrowed. |
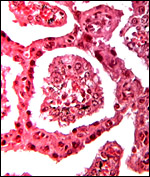 |
Higher magnification of flat section through villi interdigitating with endometrium. |
The umbilical cord of my immature specimen measured 8 cm in length, was not spiraled and had numerous plaques on its surface. The cord contains four vessels, 2 arteries and 2 veins. The veins fuse near the fetal surface. There are also numerous tiny blood vessels, as is common in artiodactyla and other orders. There is an allantoic duct that is lined with flat epithelium. Hippomanes were absent in my specimen and those described by Hradecky (1983). The plaques on the cord's surface are composed of keratinizing squamous epithelium. The remainder of the cord is covered by thin amnionic epithelium.
7) Uteroplacental circulation
This has not been described.
8)
Extraplacental membranes
There is a flat allantoic sac; yolks sacs have not been found. The amnion
is thin, avascular and pressed against the chorion and, in places, against
the allantois.
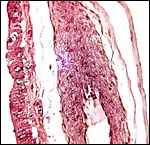 |
Allanto-chorionic membrane. Chorion at left and the thin amnion is at right. The center shows a large allantoic blood vessel. |
There is no invasion of the endometrium by trophoblast.
10) Endometrium
Steenbok have a bicornuate uterus with four distinct rows of caruncles in the endometrium. Hradecky et al. (1988) depicted an endometrial caruncle of a steenbok similar to that shown below and indicated that the endometrial glands only reach half-way up on the edematous structure. On the dome of these caruncles, the cotyledon of the placenta implants as is shown in their figure 7.
The endometrium shown below is from a post partum uterus and contains numerous hemosiderin-laden macrophages in the stroma to deep in between the large basal blood vessels. The stroma is dense connective tissue without glands, except at the edges of the caruncle. Here the epithelium has large clear cytoplasmic vacuoles.
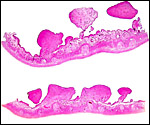 |
Sections of nonpregnant steenbok uterus showing endometrial caruncles. |
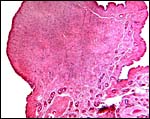 |
One of the endometrial caruncles from a post partum uterus. |
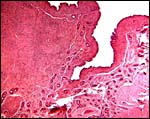 |
Margin of post partum caruncle with endometrial glands lining the lateral edge. |
None.
12) Endocrinology
I am not aware of any endocrine studies done in this species.
Steenboks have 30 chromosomes (Wallace & Fairall, 1967). All autosomes are metacentrics; the X-chromosome is a large acrocentric element and the Y is a small metacentric. Hybrids have not been described.
 |
Karyotype of male steenbok. |
 |
Karyotype of female steenbok. |
I am not aware of any publications.
15) Pathological features
Few data on pathology of steenbok are available. Cryer (1985) described
cerebrocortical necrosis. Trichostrongylus auriculatus was recovered
from a steenbok in the Kalahari National Park by Boomker (1986).
16) Physiologic data
Hay & v. Hoven (1988) showed a markedly depressive effect when tannic
acid was added to the diet of steenbok. The absorption and secretion of
fluids, electrolytes and organic acids was studied in steenbok and gerenuk
by Clemens & Moloiy (1984). They related the absorption to diets.
Hofmeyr (1981) showed that haloperidol was an effective immobilizing agent
when given intravenously. Gerneke & Cohen (1978 a,b) described infra-orbital
apocrine glands and their secretion as well as newly-described intermandibular
glands. These glands may produce secretions used in communication.
17) Other resources
CRES
at San Diego Zoo has several fibroblast cell strains that can be obtained
by contacting Dr. Oliver Ryder at oryder@ucsd.edu.
18) Other remarks - What additional
Information is needed?
Young implantation stages and endocrinology studies are needed.
Acknowledgement
The photographs come through the kindness of Mr. R. Cisneros at San Diego
Zoo.
References
Boomker, J.: Trichostrongylus auriculatus n. sp. (Nematoda: Trichostrongylidae)
from the steenbok, Raphicerus campestris (Thunberg, 1811). Onderstepoort
J. Vet. Med. 53:213-215, 1986.
Clemens, E.T. and Moloiy, G.M.: Colonic absorption and secretion of fluids, electrolytes and organic acids in East African wild ruminants. Comp. Biochem. Physiol. A 77:51-56, 1984.
Cryer, D.: Suspected cerebrocortical necrosis in the steenbok. Vet. Rec. 117:474, 1985.
Dasmann, R.F. and Mossman, A.S.: Reproduction in some ungulates in Southern Rhodesia. J. Mammal. 43:533-537, 1962.
Gerneke, W.H. and Cohen, M.: The micromorphology of the glands of the infra-orbital cutaneous sinus of the steenbok (Raphicerus campestris). Onderstepoort J. Vet. Res. 45:59-66, 1978a.
Gerneke, W.H. and Cohen, M.: The micromorphology of the apocrine glands of the intermandibular region of the steenbok (Raphicerus campestris). Onderstepoort J. Vet. Res. 45:67-74, 1978b.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Hay, L. and van Hoven, W.: Tannins and digestibility in the steenbok (Raphicerus campestris). Comp. Biochem. Physiol. A 91:509-511, 1988.
Hofmeyr, J.M.: The use of haloperidol as a long-acting neuroleptic in game capture operations. J. S. Afr. Vet. Assoc. 52:273-282, 1981.
Hradecky, P.: Placental morphology in African antelopes and giraffes. Theriogenology 20:725-734, 1983.
Hradecky, P., Mossman, H.W. and Stott, G.G.: Comparative development of ruminant placentomes. Theriogenology 29:715-729, 1988.
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Kellas, L.M.: The placenta and foetal membranes of the antelope Ourebia ourebi (Zimmermann). Acta Anat. 64:390-445, 1966.
Mentis, M.T.: A review of some life history features of the large herbivores of Africa. The Lammergeyer 16 June, 1-89, 1972.
Nowak, R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press, Baltimore, 1999.
Vrba, E.S. and Schaller, G.B.: Antelopes, Deer, and Relatives. Fossil Record, Behavioral Ecology, Systematics, and Conservation. Yale University Press, New Haven, 2000.
Wallace,
C. and Fairall, N.: The chromosomes of the steenbok. S. Afr. J. Med. Sci.
32:55-57, 1967.