 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
Saimiri sciureus
Order: Primates
Family: Cebidae
1) General Zoological Data
The squirrel monkey is widely used in research and has been classified as comprising 5 separate species by Wilson & Reeder (1992). Numerous subspecies or different names have been employed for this animal whose phenotype can differ appreciably between animals from different regions of the Amazon basin. There is also controversy, as some authors have comprised all squirrel monkeys in one species. The variability of karyotypes alone (deBoer, 1974) argues against such simplification, however. A more recent splitting of the group has been provided by Groves (2001) who gives detailed description of their phenotypes and distribution. Gotch (1979) stated that the name "Saimiri" derives from the Brazilian/Portuguese for small monkey.
This small South American cebid monkey weighs up to 1,000 g, is arboreal prefers insects, small reptiles and birds as well as fruit (see Lima & Ferrari, 2003), and is a diurnal animal. It associates in relatively large groups (over 100 animals) in the tropical rainforest and rarely comes to the floor of the forest. It is endowed with one of the largest brains for body size in primates (1/9th is CNS) and has been widely used in numerous experimental protocols. It has been useful especially in neural studies and for research into parasitology and virology. Many Primate Research Centers employ this anima for research, and so do many private laboratories. There is a huge literature on the results of many biological studies conducted in this species that has seen marked reduction in the wild and some subspecies of which are endangered now. Hof et al. (2002) who wrote about CNS aging of primates suggested that squirrel monkeys live more than 25 years.
 |
Squirrel monkey at San Diego Zoo. |
 |
Squirrel monkey at San Diego Zoo. |
Gestation lasts from 146-175 days and produces generally a single young. Twins are very uncommon. The newborn of this gestation weighed 140 g, but Hayssen et al. (1993) mentioned size up to 247 g.
3) Implantation
Dukelow et al. (1983) spent much effort to define the growth of preimplantation oocytes growth and effects on cleavage if in vitro fertilization after superovulation (16 h post hCG). They found no significantly increased aneuploidy as result of their manipulations. I have been unable to find publications on early implantation in this widely used species.
4) General Characterization of the Placenta
I have had only one delivered placenta available from a term infant that died neonatally. In addition, slides of a serially-sectioned implanted placenta were reviewed. The two placental disks, characteristic for this species and other cebidae, weighed 22 g and measured 5x4x0.5 cm each. Their surface vasculature was connected around the lateral aspect of the free membranes. This is a trabecular/villous placenta with hemochorial relationship to the maternal circulation. While Mossman (1987) mentioned the existence of villous hematopoiesis, none was found in the specimens I examined. The disks are infiltrated by a few large maternal arteries that have stiff walls, infiltrated by fibrinoid and are surrounded by syncytium (Luckett, 1974; Gruenwald, 1972).
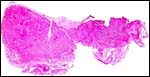
 |
Complete lobe of squirrel monkey placenta with membrane carrying connecting vessel at right. |
 |
The other disk of the same placenta, slightly thicker. Amnion at top. |
The slender villi are covered by syncytiotrophoblast as seen in the next photograph. While there is cytotrophoblast below the syncytium, at term it is so delicate that it cannot be demonstrated.
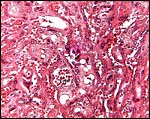 |
Trabecular villous tissue of term squirrel monkey placenta. The large amount of closely placed syncytium is obvious. |
The cord inserts near the center of one of the two disks. While this disk is often larger than the second one, that was not the case in this placenta. The cord had no twists and possessed two arteries, one vein and no ducts. It is covered by a very thin amnion. The length of umbilical cords in Saimiri was given as 8.5 cm by Spatz (1968).
7) Uteroplacental circulation
I am not aware of any fundamental research into the uterine vasculature
of this species, other than the descriptions of flow to the placenta shown
above.
The free membranes connecting the disks lack atrophied villi, unlike the condition in the human placenta. This, however, has been the case in most bilobed primate placentas. It carries the connecting vessels laterally between the two disks. The membranes have an inner single layer of epithelium without squamous metaplasia that is planted on a thin layer of connective tissue. It is loosely attached to the chorionic membrane that carries the blood vessels. Outside of the chorion is a small layer of cytotrophoblast and a very minimal amount of decidua capsularis. There are no vitelline or allantoic remnants.
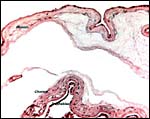 |
Membranes between lobes. |
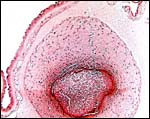 |
Interconnecting blood vessel in the chorion between the two lobes. |
Cytotrophoblast, equivalent to the "extravillous trophoblast" of human gestations superficially invades the decidua beneath the two disks and is also found to alter the maternal arterioles in a manner very similar to human placental sites. Deeper invasion does not occur.
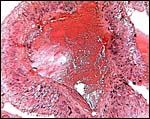 |
Maternal spiral arteriole in decidua basalis. It shows infiltration of trophoblast with partial replacement of its wall. |
There is complete decidual transformation of the endometrium and relatively few extravillous trophoblastic cells can be found deep in the endometrium. At the site of connection to the endometrium, numerous vacuolated extravillous trophoblastic cells are present intermixed with a small amount of fibrinoid.
Squirrel monkeys have no menstruation; instead, there is a well-defined estrus cycle. Ovaries have a large amount of interstitial gland tissue (Mossman & Duke, 1973).
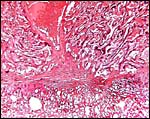 |
The large central maternal artery referred to earlier is seen in the center above the site of implantation. |
There is no subplacenta. The absence of atrophied villi in the free membranes remains unexplained.
12) Endocrinology
Wolf et al. (1977) studied the reproductive cycles and Tullner (1972)
reviewed pregnancy hormones. Positive gonadotropin tests were obtained
from 3-16 weeks of gestation. Urinary gonadotropins were positive for
7-8 weeks (see also Hodgen, 1979). Ainsworth et al. (1969) incubated squirrel
monkey placental tissue with precursors and identified production of estrone
and estradiol from androstenedione precursors and conversion of pregnenolone
to progesterone. Davidson et al. (1979) described the exceptional seasonality
of reproductive behavior in squirrel monkeys and the reaction of male
testosterone levels on females.
13)
Genetics
Squirrel monkeys have 44 chromosomes, but their fine details differ in
animals from different regions. Thus, in the karyotypes shown below (Hsu
& Benirschke, 1973), the male with 12 acrocentrics came from Leticia
(Columbia), while the female with 10 acrocentrics was from Iquitos (Peru).
Other variations have been delineated by deBoer (1974) and affect mostly
the position of centromeres, i.e. the number of acrocentrics. Stanyon
et al. (2000) who examined platyrrhines with chromosome paints suggested
that many of the former banding interpretations and putative homologies
to human chromosomes have to be re-evaluated. The same investigators (Neusser
et al., 2001) later linked the squirrel monkey as a sister group to callithricids,
as they share one derived syntenic association.
Hybrids have not been described to my knowledge and no direct crosses
between the different cytotypes are known to me. It had once been speculated
that the endangered Panamanian species (Saimiri oerstedii) might
have been introduced or be a hybrid. The study by Cropp & Boinski
(2000) has excluded this speculation.
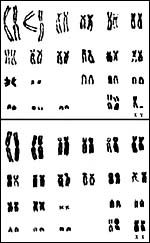 |
Male and female karyotypes of animals from different regions (see text) and having different numbers of acrocentric. |
It is likely that such studies have been performed in Primate Research Centers, but I have not been able to access fundamental information.
15) Pathological features
The most important pathologic feature of squirrel monkeys undoubtedly
is that all animals are infected with the Herpesvirus saimiri. This virus
is not pathogenic for squirrel monkeys but, as was early recognized (Hunt
et al., 1970), it causes usually fatal T-cell lymphoma in other primates,
especially marmosets. Therefore, these animals should not be housed together.
A detailed review of the genome and all other aspects of this virus are
available from Fickenscher & Fleckenstein (2001). More recently, this
virus is being considered as a potential agent for gene delivery into
cells (Whitehouse, 2003).
Scott (1992) referred to a gastric carcinoma in a squirrel monkey; also
a breast cancer was found in a male. Borda et al. (2000) described that
35% of Bolivian squirrel monkeys suffer a proliferative glomerulonephritis
with deposits of immunoglobulins. Several squirrel monkeys were found
to suffer an infectious gastritis, but Helicobacter pylori was
not found (Khanolkar-Gaitonde et al., 2000).
Many animals are infected with a variety of trypanosome, including T.
cruzi, and Ndao et al. (2000) caution handlers of New World monkeys
to be cognizant of these parasites, as they can easily be infected by
handling them or their products. Horizontal transmission of the lethal
infection with Toxoplasma gondii was clearly demonstrated by
Furuta et al. (2001). Squirrel monkeys have often been used for the study
of malaria and with hopes of vaccine production. While infection may be
achieved with some plasmodial species, they generally do not produce infectious
gametocytes. More recently, Sullivan et al. (2003) showed that infection
with Plasmodium inui shortti can infect the animals and produce
gametocytes as well.
16) Physiologic data
Numerous physiological data are available. Thus, the diet of free-ranging
monkeys was studied by Lima & Ferrari (2003); allelic frequency for
x-linked color vision was studied in two species by Cropp et al. (2002);
ABO blood groups were described by Corvelo et al. (2002). Squirrel monkeys
exhibited group A (# 67) and AB (# 7). Cortisol secretion was determined
by sampling saliva in chewed string of unrestrained squirrel monkeys by
Tiefenbacher et al. (2003). They found the expected rise after SCTH and
also determined normal daily rhythms. Numerous parameters of physiological
importance (size, life span, hematology, seminal volume etc.) have been
summarized and compared with other taxa in a table by Mather & Rushmer
(1979).
17) Other resources
Cell lines were established by Scammell et al. (2001) and, later (2002)
the same authors made a renal cell line because of the paucity of such
material available for research. But the line was hyperdiploid (2n=52-57).
18) Other remarks - What additional
Information is needed?
Descriptions of early implantation are virtually absent.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society
of San Diego.
References
Ainsworth, L., Daenen, M. and Ryan, K.J.: Steroid transformations by endocrine
organs from pregnant mammals. IV. Biosynthesis and metabolism of estrogens
and progesterone by primate placental preparation in vitro. Endocrinology
84:1421-1429, 1969.
deBoer, L.E.M.: Cytotaxonomy of the Platyrrhini (Primates). Genen Phaenen 17:1-115, 1974.
Borda, J.T., Idiart, J.R. and Negrette, M.S.: Glomerular lesions in renal biopsies of Saimiri boliviensis (primate) examined by light and electron microscopy and immunohistochemistry. Vet. Pathol. 37:409-414, 2000.
Corvelo, T.C., Schneider, H. and Harada, M.L.: ABO blood groups in the primate species of Cebidae from the Amazon region. J. Med. Primatol. 31:136-141, 2002.
Cropp, S. and Boinski, S.: The Central American squirrel monkey (Saimiri oerstedii): Introduced or endemic species? Molec. Phylogenet. Evol. 16:350-365, 2000.
Cropp, S., Boinski, S. and Li, W.H.: Allelic variation in the squirrel monkey x-linked color vision gene: biogeographical and behavioral correlates. J. Mol. Evol. 54:734-745, 2002.
Davidson, J.M., Gray, G.D. and Smith, E.R.: Animal models in the endocrinology of reproductive behavior. Chapter 5 (pp. 61-81) in, Animal Models for Research on Contraception and Fertility, N.J. Alexander, ed. Harper & Row, Hagerstown, 1979.
Dukelow, W.R., Chan, P.J., Hutz, R.J., Demayo, F.J., Dooley, V.D., Rawlins, R.G. and Ridha, M.T.: Preimplantation development of the primate embryo after in vitro fertilization. J. Exp. Zool. 228:215-221, 1983.
Fickenscher, H. and Fleckenstein, B.: Philos. Trans. R. Soc. B Biol. Sci. 356:545-567, 2001.
Furuta, T., Une, Y., Omura, M., Matsutani, N., Nomura, Y., Kikuchi, T., Hattori, S. and Yoshikawa, Y.: Horizontal transmission of Toxoplasma gondii in squirrel monkeys (Saimiri sciureus). Exp. Anim. 50:299-306, 2001.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Groves, C.: Primate Taxonomy. Smithsonian Institution Press, Washington, 2001.
Gruenwald, P.: Expansion of placental site and maternal blood supply of primate placentas. Anat. Rec. 173:189-204, 1972.
Hayssen, V., van Tienhoven, A. and van
Tienhoven, A.: Asdell's Patterns of Mammalian Reproduction: a Compendium
of Species-specific Data. Comstock/Cornell University Press, Ithaca, 1993.
Hodgen, G.D.: Primate models for pregnancy hormone secretion in man: fetal,
maternal, and placental factors. Chapter 32 (pp. 425-436) in, Animal Models
for Research on Contraception and Fertility, N.J. Alexander, ed. Harper
& Row, Hagerstown, 1979.
Hof, P.R., Gilissen, E.P., Sherwood, C.C., Duan, H., Lee, P.W.H., Delman, B.N., Naidich, T.P., Gannon, P.J., Perl, D.P. and Erwin, J.M.: Comparative neuropathology of brain aging in primates. Pp. 130-154, in, Aging in Nonhuman Primates, J.M. Erwin, ed. Interdisciplinary Topics in Gerontology, Vol. 31, Karger, Basel, 2002.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 7, Folio 348, 1973. Springer-Verlag, NY.
Hunt, R.D., Melendez, L.V., King, N.W., Gilmore, C.E., Daniel, M.D., Williamson, M.E. and Jones, T.C.: Morphology of a disease with features of malignant lymphoma in marmosets and owl monkeys inoculated with Herpesvirus saimiri. J. Natl. Cancer Inst. 44:447-465, 1970.
Khanolkar-Gaitonde, S.S., Reubish, G.K., Lee, C.K. and Stadtlander, C.T.: Isolation of bacteria other than Helicobacter pylori from stomachs of squirrel monkeys (Saimiri spp.) with gastritis. Dig. Dis. Sci. 45:272-280, 2000.
Lima, E.M. and Ferrari, S.F.: Diet of a free-ranging group of squirrel monkeys (Saimiri sciureus) in eastern Brazilian Amazonia. Fol. Primatol. 74:150-158, 2003.
Luckett, W.P.: Comparative development and evolution in the placenta in primates. In: Contributions in Primatology, Vol. 3, W.P. Luckett ed., pp.142-234, 1974. S. Karger, Basel.
Mather, E.C. and Rushmer, R.A.: Physiological
parameters of some species used in reproduction research. Pp. 561-576,
in, Animal Models for Research on Contraception and Fertility, N.J. Alexander,
ed. Harper & Row, Hagerstown, 1979.
Mossman, H.W.: Vertebrate Fetal Membranes. MacMillan, Houndmills, 1987.
Scott, G.B.D.: Comparative Primate Pathology. Oxford University Press,
1992.
Mossman, H.W. and Duke, K.L.: Comparative Morphology of the Mammalian Ovary. University of Wisconsin Press, Madison, Wisconsin, 1973.
Ndao, M., Kelly, N., Normandin, D., Maclean, J.D., Whiteman, A., Kokoskin, E., Arevalo, I. and Ward, B.J.: Trypanosoma cruzi infection in squirrel monkeys: comparison of blood smear examination, commercial enzyme-linked immunosorbent assay, and polymerase chain reaction analysis as screening tests for evaluation of monkey-related injuries. Comp. Med. 50:658-665, 2000.
Neusser, M., Stanyon, R., Bigoni, F., Wienberg, J. and Muller, S.: Molecular cytotaxonomy of New World monkeys (Platyrrhini) - comparative analysis of five species by multi-color chromosome painting gives evidence for classification of Callimico goeldii within the family of Callitrichidae. Cytogenet. Cell Genet. 94:206-215, 2001.
Scammell, J.G., Wright, J.L. and Tuck-Muller, C.M.: The origin of four squirrel monkey cell lines established by karyotype analysis. Cytogenet. Cell Genet. 93:263-264, 2001.
Scammell, J.G., Tucker, J.A., King, J.A., Moore, C.M., Wright, J.L. and Tuck-Muller, C.M.: A kidney epithelial cell line from a Bolivian squirrel monkey. In Vitro Cell Devel. Biol. Anim. 38:258-261, 2002.
Spatz, W.B.: Nabelschnur-Längen bei Insektivoren und Primaten. Z. Säugetierk. 33:226-239, 1968.
Stanyon, R., Consigliere, S., Muller, S., Morescalchi, A., Neusser, M. and Wienberg, J.: Fluorescence in situ hybridization (FISH) maps chromosomal homologies between the dusky titi and squirrel monkey. Amer. J. Primatol. 50:95-107, 2000.
Sullivan, J.S., Nace, D., Williams, T., Guarner, J., Noland, G.S. and Collins, W.E.: The development of exoerythrocytic stages of Plasmodium inui shortti in New World monkeys. J. Parasitol. 89:637-639, 2003.
Tiefenbacher, S., Lee, B., Meyer, J.S. and Spealman, R.D.: Noninvasive technique for the repeated sampling of salivary free cortisol in awake, unrestrained squirrel monkeys. Amer. J. Primatol. 60:69-75, 2003.
Tullner, W.W.: Chorionic gonadotrophin in non-human primates. Pp. 200-213, in, WHO Symposium on the Use of Non-Human Primates for Research on Problems of Human Reproduction. E. Diczfalusy & C.C. Standley, eds., 1972. Also as: Acta Endocrinol. Suppl. 166.
Whitehouse, A.: Herpesvirus saimiri: a potential gene delivery vector (review). Int. J. Mol. Med. 11:139-148, 2003.
Wilson, D.E. and Reeder, D.A.M.: Mammal Species of the World. A Taxonomic and Geographic Reference. 2nd ed. Smithsonian Institution Press, Washington, DC, 1992.
Wolf, R.C., O'Connor, R.F. and Robinson, J.A.: Cyclic changes in plasma progestins and estrogens in squirrel monkeys. Biol. Reprod. 17*:228-231, 1977.