 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last updated: May 4, 2007. |
Gazella spekei
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
Speke's gazelles come from Eastern Ethiopia and Northern Somalia and are currently endangered with their numbers having been drastically reduced recently. Only slightly more than 1,000 animals are currently thought to live in the wild. Their longevity is 12 years 8 ms. (Jones, 1993). The major colony in this country exists at the St. Louis Zoological Park that started a breeding program in 1972 with 1.3 animals See Read & Frueh, 1980). Many inbreeding studies have since been done on that colony's success ( v.i. ). The nose is this species' most characteristic feature, as the upper dark portions may be voluntarily inflated considerably, to the size of a tennis ball. It allows the production of an alarm call that is said to sound like a shot. The inflated nose is beautifully shown in a photograph by Read & Frueh (1980).
 |
Adult Speke's gazelle at San Diego Zoo. |
2) General Gestational Data
The usually single newborns weigh ~1,250 g (Nowak, 1,999). Read & Frueh (1980 recorded weights in females from 1,344 to 1,792 g, and from 1,344 to 1,792 g in males. The latter authors indicate that first conception occurs at 16.8 months. They indicate that the length of gestation is 169-190 days. The adult females weigh from 11-16 kg; males weigh 18 kg.
3) Implantation
There are no such studies.
4) General Characterization of the Placenta
The multicotyledonary placenta that I had available weighed 190 g and had 80 cotyledons that measured between 2.5 and 1 cm in width and 1 cm in depth. It must have extended into both uterine horns. In April, 2007 a dam died from renal neoplasms whilst pregnant. A gradual distension of the abdomen had been noted and a large amount of peritoneal fluid was found at autopsy. The female fetus weighed only 350 g, and the uterus with placenta weighed 350 g. There were 47 cotyledons located in primarily the left horn but extending into the right horn as well. The right, unoccupied horn had much smaller cotyledons. The largest cotyledon measured 4 x 2.5 cm. The umbilical cord measured 6 x 1.2 cm and contained 4 vessels and had fine granules of squamous metaplasia covering its surface. It contained a large allantoic duct and had numerous small vessels in Wharton’s jelly.
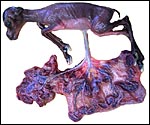 |
Fetus in left uterine horn with cotyledons expanding into both horns. |
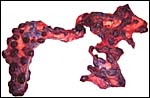 |
Term placenta with 80 cotyledons. |
5) Details of fetal/maternal barrier
The villous structure is similar to that of other bovid species. The fetal/maternal relation is epitheliochorial. Occasional binucleate cells interrupt the single-layered cuboidal trophoblast of the villous surface. The fetal capillaries protrude frequently into the trophoblastic cover of the villi and seem then to replace the trophoblast. No pigment of the hemophagous kind was present in this first placenta, however, there was extensive pigmentation in the trophoblast beneath the chorionic surface.
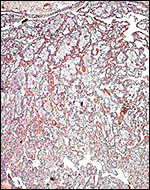 |
Low power view of villous structures with chorion above. |
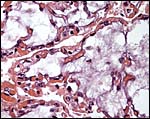 |
Many villi are not only showing autolysis (myxoid core) but also prominent surface capillaries. |
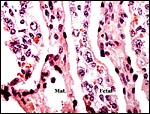 |
Interspersion of villi (Fetal) with maternal (Mat.) parenchyma. |
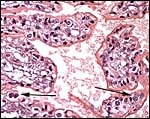 |
Higher magnification of trophoblastic surface with binucleate cells at arrow. |
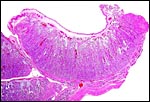 |
Implanted small cotyledons in the lesser (right) horn. |
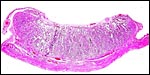 |
Implanted cotyledon of second case. |
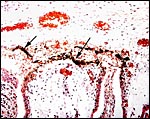 |
Surface of the placenta with arrows pointing to pigmented trophoblast. |
6) Umbilical cord
The umbilical cord measured 15 cm in length, had no spirals and contained 4 vessels and the allantoic duct. Its surface was smooth. The surface of the 6 cm long umbilical cord from the stillborn fetus shown above had fine granules of squamous metaplasia.
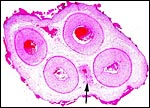 |
Cross section of umbilical cord of case two with arrow indicating the allantoic duct. Note the numerous areas of squamous metaplasia on the surface. |
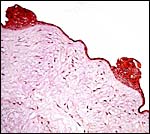 |
Surface of umbilical cord with numerous areas of squamous metaplasia. |
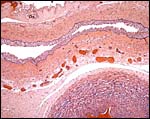 |
Allantoic duct in the umbilical cord. Note the extensive small vessel presence. |
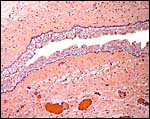 |
Higher magnification of allantoic duct epithelium. |
7) Uteroplacental circulation
This has not been studied.
8) Extraplacental membranes
The amnion is thin and avascular. It has a single layer of thin squamous epithelium without keratinization. The large, vascularized allantoic sac is covered by a thick epithelium. No decidual transformation takes place on the outside of the membranes.
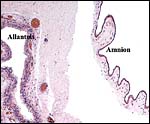 |
Membrane composition of amnion at right and vascularized allantoic sac at left. |
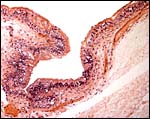 |
Higher magnification of allantoic membrane epithelium. |
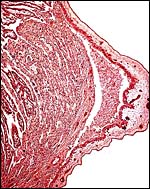 |
Edge of small cotyledon with origin of intercotyledonary membranes. |
9) Trophoblast external to barrier
There is none.
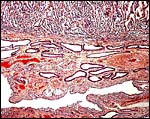 |
Implanted placenta of second case to show ovoid endometrial glands and absence of trophoblastic invasion. |
10) Endometrium
True decidual transformation of the endometrium does not occur and no maternal tissue accompanies the delivered placenta.
11) Various features
There is no subplacenta.
12) Endocrinology
Progesterone levels were used to determine pregnancy by Read & Frueh (1980), but they did not provide data. In addition, they used sonography and radiology to ascertain gestations.
13) Genetics
Males have 33 chromosomes, females have 32 (Hsu & Benirschke, 1974). They are very similar to those of Dorcas gazelles; there is a large X/A fusion chromosome with the distal portion of the compound X being the autosome. The putative evolution of the composite X chromosome was studied by Effron et al. (1976). It is diagrammed below. Hybrids have not been described.
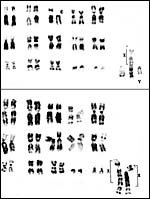 |
Typical chromosome sets of male and female Speke's gazelles. |
 |
Hypothetical evolution of compound X-chromosome (Effron et al., 1976). |
The small founder population of Speke's gazelle at St. Louis zoo (1.3 animals in 1972) and the variable success in reproduction (hundreds of animals by now) has been studied numerous times by population geneticists but there is no unanimity which factors are responsible for the reproductive successes. In 1982 there were 44 living animals in the USA and MacCluer et al. (1986) analyzed the gene loss (mostly of offspring from one female) in the population. In 2000 there were 41 animals in US zoos. Elective inbreeding with dropping of genes, the effect of different housing in various zoological gardens and their respective conditions, and other possibilities for the results have been discussed at length (Kalinowski et al., 2000; Templeton & Read, 1983, 1998; Willis & Wiese, 1997). Suffice it to say here that a healthy, large population of gazelles now exists and that congenital abnormalities (as possibly expected from inbreeding) are not apparent. New stock was also introduced in subsequent years and now several zoos breed this species. The St. Louis Zoo is responsible for the studbook (Fischer, 1993). Butler et al. (1994) described results from DNA fingerprints and thus identified new genetic contributions to the stock at St. Louis . By measuring intraerythrocytic K and Na concentrations, Georgiadis et al. (1988) were able to demonstrate HK/LK polymorphism.
14) Immunology
I am not aware of any data.
15) Pathological features
Despite the large number of gazellinae kept at San Diego Zoo, Griner (1983) did not list this species as having been autopsied. Boever & Herbers (1977) observed a congenital sebaceous adenoma, and Munson & Miller (1986) felt that this species is prone to develop hepatic abscesses. Hypophosphatemia and hypovitaminosis-D with rib fractures were reported in young gazelles by Weber & Junge (2001). Iori & Lanfranchi (1996) studied the endoparasites in Speke's gazelles of Somalia . Read & Frueh (1980) experienced a high neonatal mortality (16 of 54 births) and performed Cesarean sections with stillborn calves. Fives neonates suffered a patent ductus arteriosus, injury by cagemates killed thee animals, and septicemia two others. Pneumonia occurred in aged animals, enterotoxemia' took place after dietary changes, but trauma (legs and teeth) were also prominent causes of death. Strongyloides and coccidian infections responded to therapy. The dam whose female fetus was shown above had died from bilateral renal neoplasms shown next. Subsequently the right uterine horn is shown that had endometritis/salpingitis and hemosiderin-laden macrophages, presumably leftovers from an earlier gestation.
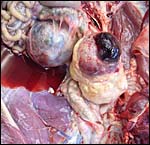 |
Bilateral renal neoplasms of the dam. |
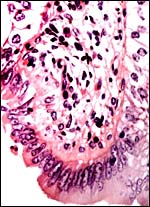 |
Right uterine horn showing chronic inflammation. |
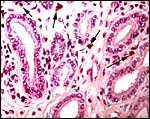 |
Hemosiderin-laden macrophages in same horn. |
16) Physiologic data
I am not aware of any data other than the experimentation in St. Louis with artificial insemination. This was done after prostaglandin and GnRH-releasing hormone injections. Fresh artificially ejaculated and also frozen semen was used in three animals successfully with gestations resulting with gestational ages of 177, 189 and 180 days respectively.
17) Other resources
Cell lines are available from CRES at San Diego Zoo.
18) Other remarks What additional Information is needed?
Early implantation needs to be studied.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society of San Diego.
References
Boever, W.J. and Herbers, H.: Congenital sebaceous gland adenoma in a Speke's gazelle. Vet. Med. Small Anim. Clin. 72:268-269, 1977.
Butler , M.A., Templeton, A.R. and Read, B.: DNA fingerprinting in Speke's gazelle: a test for genetic distinctness, and the correlation between retardedness and similarity. Mol. Evol. 3:355-361, 1994.
Effron, M., Bogart, M.H., Kumamoto , A.T. and Benirschke, K.: Chromosome studies in the mammalian subfamily Antilopinae . Genetica 46: 419-444, 1976.
Fisher, M.: Speke's gazelle studbook ( Gazella spekei ). St. Louis Zoological Park, St. Louis , Missouri , 1993. (fischer@stlzoo.org)
Georgiadis, N., Dunham, P., Read, B. and Templeton, A.: HK/LK polymorphism and its genetic determination in Speke's gazelle. J. Hered. 79:325-331, 1988.
Griner, L.A. : Pathology of Zoo Animals. Zoological Society of San Diego , San Diego , California , 1983.
Hsu, T.C. and Benirschke, K.: Atlas of Mammalian Chromosomes 8: Folio 395, 1974.
Iori, A. and Lanfranchi, P.: Contribution to the knowledge of helminthofauna of wild mammals of Somali. Parassitologia 38:511-515, 1996.
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Kalinowski, S.T., Hedrick, P.W. and Miller, P.S.: Inbreeding depression in the Speke's gazelle captive breeding program. Conserv. Biol. 14:1375-1384, 2000.
Munson, L. and Miller, R.E.: Susceptibility of the Speke's gazelle to hepatic abscesses. J. Amer. Vet. Med. Assoc. 189:1056-1058, 1986.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Read, B. and Frueh, R.J.J.: Management and breeding of Speke's gazelle Gazella spekei at St. Louis Zoo, with a note on artificial insemination. Intern. Zoo Ybk. 20:99-104, 1980.
Templeton, A.R. and Read, B.: The elimination of inbreeding depression in a captive herd of Speke's gazelle. In: Schoenwald-Cox, C.M., Chambers, S.M., MacBryde, M. and Thomas, L. eds. Genetics and Conversation. Benjamin/Cummings, Menlo Park , California , pp. 241-261, 1983.
Templeton, A.R. and Read, B.: Elimination of inbreeding depression from a captive population of Speke's gazelle: validity of the original statistical analysis and confirmation by permutation testing. Zoo Biol. 17:77-98, 1998.
Weber, M.A. and Junge, R.E.: Hypophosphatemia associated with hypovitaminosis-D in Speke's gazelles (Gazella spekei). J. Zoo Wildl. Med. 32:379-383, 2001.
Willis, K. and Wiese, R.J.: Elimination of inbreeding depression from captive populations: Speke's gazelle revisited. Zoo Biol. 16:9-16, 1997.
MacCluer, J.W., VandeBerg, J.L., Read, B. and Ryder, O.A.: Pedigree analysis by computer simulation. Zoo Biol. 5:147-160, 1986.