| |
9)
Trophoblast external to barrier
No implanted specimens have been studied and thus it is unknown whether
trophoblast invasion occurs. But, in view of the general similarities to
other ungulate placentations, this is unlikely.
10)
Endometrium
It is unknown whether true decidua develops, but it is unlikely.
11)
Various features
No remarkable additional features are worth describing.
12)
Endocrinology
Sempere et al. (2001) studied Persian and sand gazelles from a reproductive
cycle point of view and determined periods of anestrus and estrus. They
measured progesterone and prolactin levels in both species. Melatonin
treatment depressed prolactin levels.
13) Genetics
The cytogenetics of goitered gazelles is complex. This is probably due
to the fact that the precise place of origin of zoo-held animals is usually
unknown and, from reading the literature, it would appear that there are
hybrids among what were considered to be subspecies. Some of these animals,
those with different chromosome number, should probably be given species
designation. Since the animals extend over such a wide region (from Arabia
to Mongolia), several "hybrid zones" probably exist and there
is intergrading of different types.
Wurster (1972) found 2n=31 in two male, and 2n=30 in one female G.
subgutturosa. The uneven number of male/female animals is the result
of a translocation of an autosome to the X-chromosome. This is a common
feature of many gazelles and antelopes of Africa and seems to have originated
before the invasion of Africa by ungulates from Eurasia. It was studied
extensively by Effron et al. (1976). This karyotype for G. s. subgutturosa
was confirmed by Benirschke & Kumamoto (1987) who found, in numerous
sand gazelles (G. s. marica) studied, varied karyotypes. For females
there were 2n=32 and 30, and for males 2n=33 and 31. Hybridization between
Persian and sand gazelles was assumed to be the reason and it was deemed
not to have resulted in subfertility. Similarly, Granjon et al. (1991)
assumed hybridization, as they found male sand gazelles from Saudi Arabia
with 2n=33 and 2n=31, and females with 2n=32. Nevertheless, our females
possessed 2n=31. Kingswood et al. (1994) studied meiosis and karyotypes
of Persian gazelles with heterozygosity for the 14/15 autosomal translocation
and found that this did not impair fertility. Vassart et al. (1993) found
in males from Saudi Arabia and Qatar 33, 32, 31 chromosomes, and in females
32, 31, 30. It involved the same Robertsonian translocations. The problem
again being that the precise origin of the population was unknown. Other
studies exist as that by Vassart et al. (1995) which also includes protein
electrophoretic data. In addition, an important finding of Chinese G.
s. subgutturosa by Orlov (1987) identified 2n=30/31; this is essential
to know, as no other subspecies exist in that region.
14)
Immunology
No studies have been published.
15)
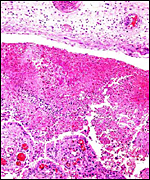
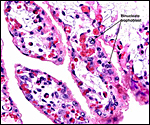
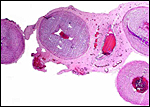
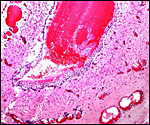
Pathological features
Mohammed & Flamand (1996) succeeded in experimental infection of Arabian
sand gazelles with Eimeria rheemi. This is a "natural"
and frequent infection of the rheem (sand gazelle) and causes diarrhea
(Hussein & Mohammed, 1992). Sand gazelles and other local Arabian
gazelles are often infected with sarcocystis parasites (Mohammed et al.,
2000). Fenwick (1983) described cryptosporidiosis in a neonatal Persian
gazelle.
Griner (1983) did not differentiate the various types of gazelles and
found that among "Persian gazelles" the primary cause of death
was trauma and, in neonates, "malnutrition".
16) Physiologic data
Hematologic data were provided for mountain gazelles (Gazella gazella)
by Rietkerk et al. (1994), while Vassart et al. (1994) gave the details
for serum chemistry values in sand gazelles. The natural diet was detailed
by Mohamed et al. (1991). A very detailed description of the behavior
of all kinds of gazelles was published by Walther (1968).
17) Other resources
The research department (CRES)
of the Zoological Society of San Diego has had a long interest in the
genetics of gazelles, and, especially in the polymorphism of goitered
gazelles (Kingswood et al., 1994). As a consequence, numerous cell strains
are available of various karyotypes by requesting them from Dr. Oliver
Ryder at oryder@ucsd.edu.
There is a very extensive bibliography on all aspects of sand gazelles
and the related forms. This has been gathered by the King Khalid Wildlife
Research Center, Thumammah and can be made available by the National Commission
for Wildlife Conservation and Development, P.O. Box 61681, Riyadh 11575,
Kingdom of Saudi Arabia.
18) Other remarks - What additional Information is needed?
Information is needed on reproductive endocrine data and the cytogenetics
of the other species of goitered gazelles.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society
of San Diego. I appreciate also very much the help of the pathologists
at the San Diego Zoo.
References
Benirschke, K. and Kumamoto, A.T.: Challenges of artiodactyl cytogenetics.
La Kromosoma II-45:1468-1478 1987.
Carter,
S. and Kingswood, S.: Goitered Gazelle. North American Regional Studbook
First edition. Sedgwick County Zoo & Botanical Garden, Wichita, Kansas,
1991.
Effron,
M., Bogart, M.H., Kumamoto, A.T. and Benirschke K.: Chromosome studies
in the mammalian subfamily Antilopinae. Genetica 46:419-444, 1976.
Fenwick,
B.W.: Cryptosporidiosis in a neonatal gazella. JAVMA 183:1331, 1983.
Granjon,
L., Vassart, M., Greth, A. and Cribiu, E.-P.: Genetic study of sand gazelles
(Gazella subgutturosa marica) from Saudi Arabia. Chromosomal and
isozymic data. Z. Säugetierk. 56:169-176, 191.
Griner,
L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego,
California, 1983.
Groves,
C.P.: An introduction to the gazelles. Chinkara: Bulletin of the Gazelle
Research Group. 1(1):4-16, 1985.
Hussein,
H.S. and Mohammed, O.B.: Eimeria rheemi sp. n. (Apicomplex: Eimeriidae)
from the Arabian Sand Gazelle, Gazella subgutturosa marica (Artiodactyla:
Bovidae) in Saudi Arabia. J. Helminthol. Soc. Wash 59:190-194, 1992.
Kingswood,
S.C., Kumamoto, A.T., Sudman, P.D., Fletcher, K.C. and Greenbaum, I.F.:
Meiosis in chromosomally heteromorphic goitered gazelle, Gazella subgutturosa
(Artiodactyla, Bovidae). Chromosome Research 2:37-46, 1994.
Mohamed,
A., Abbas, J. and Salch, M.: Natural diet of the Arabian Rheem gazelle,
Gazella subgutturosa marica. J. Arid Environm. 20:371-374, 1991.
Mohammed,
O.B. and Flamand, J.R.: Experimental infection of Arabian sand gazelles,
Gazella subgutturosa marica with Eimeria rheemi. J. Parasitol.
8:356-37, 1996.
Mohammed,
O.B., Davies, A.J., Hussein, H.S. and Daszak, P.: Sarcocystis infections
in gazelles at the King Khalid Wildlife Research Centre, Saudi Arabia.
Vet. Rec. 146:218-2, 2000.
Orlov,
in Shi, L.: Recent trends in mammalian cytogenetics in China. La Kromosoma
II-45:1458-1467, 1987.
Rietkerk,
F.E., Delima, E.C. and Mubarak, S.M.: The hematological profile of the
mountain gazelle (Gazella gazella): variations with sex, age, capture
method, season, and anesthesia. J. Wild. Dis. 30:6976, 1994.
Rostron,
J.: A multivariate statistical study the skull measurements of five taxa
of gazelles. Biol. J. Linn. Soc. 4:1-14, 1972.
Sempere,
A.J., Brown, N., Pereladova, O.B., Bahloul, K., Lacroix, A. and Soldatova,
N.: Comparative analysis of reproductive cycles in female Persian gazelle
(Gazella subgutturosa subgutturosa) (Central Asia) and sand gazelle
(Gazella subgutturosa marica) (Arabian Peninsula). Gen. Comp. Endocrinol.
121:57-65, 2001.
Vassart M, Greth A, Durand V and Cribiu EP. Chromosomal polymorphism in
sand gazelles (Gazella subgutturosa marica). J Heredity 84:478-481
, 1993.
Vassart,
M., Greth, A., de la Farge, F. and Braun, J.P.: Serum chemistry values
for Arabian sand gazelles (Gazella subgutturosa marica). J. Wildl.
Dis. 30:426-428, 1994.
Vassart,
M., Granjon, L. and Greth, A.: Genetic study of Gazella gazella:
chromosomal and allozymic data. C. R. Acad. Sc. III 318:27-33, 1995.
Walther,
F.: Die Gazellen und ihre Verwandten. In, Gzimeks Tierleben. Volume 13.
Kindler-Verlag Zürich, 1968.
Walther,
F.: Verhalten der Gazellen. Neue Brehms Bücherei, Ziemsen Verlag,
Wittenberg, 1968.
Wurster,
D.H.: Sex chromosome translocations and karyotypes in bovid tribes. Cytogenetics
11:197-207, 1972.
|