 |
|
(Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.)
|
| Last updated: April 20, 2004. |
Gazella subgutturosa subgutturosa
Order:
Artiodactyla
Family: Bovidae
1) General Zoological Data
In
contrast to most other gazelles, goitered gazelles possess horns only
in males. These gazelles inhabit desert areas from Palestine to Northern
China and have become severely reduced in numbers by hunting. A related
species, the Sand gazelle (Gazella subgutturosa marica) has previously
had a chapter (see Sand gazelle chapter, as the chapters overlap).
"The
goitered gazelle, Gazella
subgutturosa, inhabits
desert and sub-desert steppes of the Arabian peninsula and southwestern
Asia to northern China and Mongolia. Four subspecies are recognized by
Groves (1985), two of which are presently maintained in captivity in North
America. The Arabian sand gazelle (see chapter on Sand gazelle),
G.
s. marica,
was once common in the Arabian peninsula, ranging north through eastern
Jordan and Iraq, where its range integrated with that of G.
s. subgutturosa,
commonly known as the Persian or goitered gazelle. Persian gazelles once
ranged from eastern Turkey through Iran, Pakistan and Soviet Central Asia.
The two forms that are not maintained in North American collections are
the Xinjiang goitered gazelle, G. s. yarkandensis, found in the
deserts of the Tarim Basin, Xinjiang, China, and the Mongolian goitered
gazelle G. s. hilleriana, native to the Gobi and Caidam (Groves, 1985,
see Fig. 1). The only captive records of these forms are from European
collections. These records indicate that G. s. hilleriana specimens
were present in the Opel Zoo as recently as the 1970's, while four G.
s. yarkandensis were held by the Berlin Zoo from 1904 to 1906."
This information has been taken from the sand gazelle studbook report's
initial statement (Carter & Kingswood, 1991). Relatively small colonies
of sand gazelles are now held in a few zoos, both in Europe and in the
USA. In addition, it has apparently been difficult to differentiate clearly
between the different phenotypes of these gazelles by mere inspection
of the four subspecies, if this is possible at all. This is also apparent
from the two photographs shown in the chapter on sand gazelles. Perhaps
specific phenotypes have special, as yet unrecognized chromosomal characteristics
as well. I am not aware that such comparisons (karyotypes with photographs)
have ever been attempted. Rostron (1972) did a careful study of skull
measurements of Arabian gazelles and thereby assigned species designation.
 |
Male Persian gazelle at San Diego Wild Animal Park. |
Twins occur in at least one third of goitered gazelle pregnancies (Walther, 1968; Nowak, 1999). Newborns weigh on average 1,250 g.
3) Implantation
Early stages of implantation have not been observed, thus the pregnant uterus to be described presently is of interest.
4)
General Characterization of the Placenta
One singleton near-term placenta (34 cm CR male conceptus in utero) and
an additional uterus from an immature twin gestation were available. The
first uterus is from a 6 year-old dam that died after severe trauma. The
uterus and placenta were not injured. The twins of the second gestation
were 11.5 cm CR length and weighed 63.7 and 57.3 g. The placenta of each
fetus had 60 cotyledons. Each twin and its placenta were in separate uterine
horns. Each umbilical cord measured 7 x 0.7 cm and both were centrally
inserted. The cotyledons were of relatively uniform size and firmly intertwined
with the caruncles. There were approximately four somewhat irregular rows
of cotyledons/caruncles. The dam had to be euthanized during this pregnancy
because of numerous traumatic wounds she had sustained.
An additional uterus with fraternal twins that were closer to term than the previous uterus has since become available. The male weighed 750 g (25 cm CR), the female twin was 720 g (28 cm CR). The entire uterus with attached placentas weighed 750 g and there were 42 and 44 cotyledons in four rows respectively. Each umbilical artery measured 10 cm in length, had four vessels and an allantoic duct. They were covered with fine granules of squamous metaplasia.
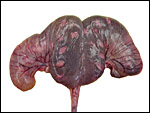 |
Bicornuate uterus with young twins in situ. |
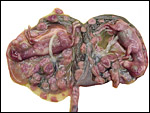 |
Opened uterus with immature twins and attached placenta. |
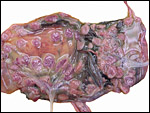 |
After removal of the fetuses, the placentas remain attached in both uterine horns. |
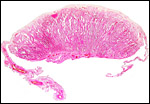 |
Complete cotyledon attached to the uterus with fetal surface above. |
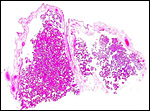 |
Microscopic picture of two adjacent cotyledons from the placenta of the first gestation. |
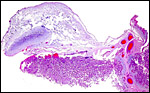 |
Another cotyledon of the near-term placenta with attached umbilical cord and intercotyledonary membranes. |
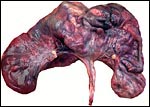 |
Second uterus with twins of Persian gazelle. The ovaries are the two small yellow corpuscles next to cervix. |
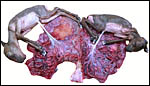 |
Previous specimen opened with male at left, female at right horn. |
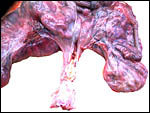 |
Cervix filled with thick yellow mucus of previous specimen. |
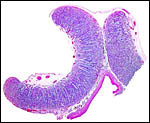 |
Two adjacent cotyledons from previously shown placenta. |
This is a typical epitheliochorial, polycotyledonary placenta with large allantoic cavity. The trophoblast is single-layered and has numerous binucleate cells. Beneath the chorionic plate the trophoblast is more cylindrical. Pigmentations of trophoblast, as seen in many other ungulates, were not present.
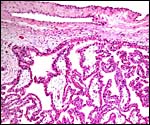 |
Fetal surface of immature cotyledon with chorion and fused amnion above. |
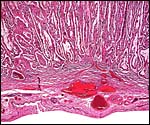 |
Maternal attachment of immature cotyledon. Few endometrial glands remain, and there is no typical decidua. The interdigitation of maternal endometrial septa with villi is readily apparent. |
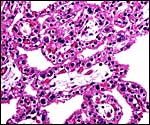 |
Immature villi of Persian gazelle with numerous binucleate cells, some at white arrows. |
The umbilical cord has four large blood vessels and numerous small allantoic vessels, especially distributed around the prominent allantoic duct. The surface of the umbilical cord has numerous foci of squamous metaplasia with keratin production in the mature placenta.
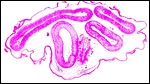 |
Cross section of mature umbilical cord with allantoic duct in the center below. A few foci of squamous metaplasia are on the surface. |
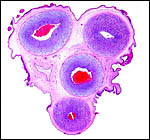 |
Cord of one twin with allantoic duct at left and numerous squamous plaques on surface. |
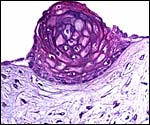 |
Focus of squamous metaplasia on the cord surface. |
This has not been studied.
8)
Extraplacental membranes
The amnion has a flat squamous epithelium and applies closely to the allantoic
membrane. The latter has columnar epithelium and is vascularized. There
is no real decidua developed in the uterus. Hippomanes have not been seen.
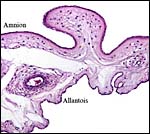 |
Amnion and allantoic membranes. |
No trophoblastic invasion into the endometrium occurs.
10)
Endometrium
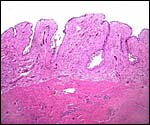
Whether a true decidua develops is unknown but the study of this specimen
suggests that there is no real decidualization. In the immature fetal
uterus, the development of caruncles is obvious.
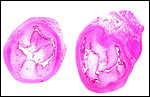 |
Cross sections through the uterine horns of one immature fetus showing the endometrial caruncles. |
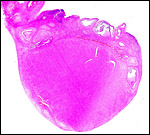
 |
Neonatal uterus with endometrial caruncles. |
There is no subplacenta and no metrial glands develop.
12)
Endocrinology
Sempere et al. (2001) studied reproductive cycles of Persian and sand
gazelles and determined periods of anestrus and estrus. They measured
progesterone and prolactin levels in both species. Melatonin treatment
depressed prolactin levels.
 |
Corpus luteum and Graafian follicle development in the immature gestation. |
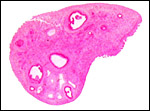 |
Immature neonate ovary with follicular development (2 ms.). |
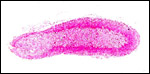 |
Adrenal gland of 2 months-old neonate. |
Persian gazelles have the characteristic X-autosome fusion that is shared by many other gazelles. They thus possess 31 chromosomes in the male animal and 30 chromosomes in female (Wurster, 1972; Effron et al., 1976; Hsu & Benirschke, 1977). Hybrids of Persian gazelles with Marica gazelles (Gazella leptoceros marica) occurred in the London Zoo (Gray1972). The X/A fusion is a common feature of many gazelles and antelopes of Africa and it seems to have originated before the invasion of Africa by ungulates from Eurasia. It was studied extensively by Effron et al. (1976). This karyotype of G. s. subgutturosa was confirmed by Benirschke & Kumamoto (1987) who found, in numerous sand gazelles (G. s. marica) studied, varied karyotypes. For females there were 2n=32 and 30, and for males 2n=33 and 31. Hybridization between Persian and sand gazelles was assumed to be the reason for this admixture and it was deemed not to have resulted in subfertility. Similarly, Granjon et al. (1991) assumed hybridization, as they found male sand gazelles from Saudi Arabia with 2n=33 and 2n=31, and females with 2n=32. Nevertheless, our females possessed 2n=31. Kingswood et al. (1994) studied meiosis and karyotypes of Persian gazelles with heterozygosity for the 14/15 autosomal translocation and found that this did not impair fertility. Vassart et al. (1993) found in males from Saudi Arabia and Qatar 33, 32, 31 chromosomes, and in females 32, 31, 30. The process involved the same Robertsonian translocations, with the same autosome. The problem of correct species assignment was again apparent, with the precise place of origin of the population remaining unknown. Other studies exist, as that by Vassart et al. (1995), which also includes protein electrophoretic data. In addition, an important finding is the study of the Chinese G. s. subgutturosa (Orlov, 1987) with identification of 2n=30/31 chromosome numbers. This is essential to know, as no other subspecies exist in that region and hybridization can presumably be excluded.
 |
Male and female karyotypes of Persian gazelles. |
I am not aware of any studies in this field.
15)
Pathological features
Griner (1983) did not differentiate the various types of gazelles and
found that, among "Persian gazelles", the primary cause of death
was trauma and neonates had "malnutrition" as major problems..
16)
Physiologic data
Some data (hematologic and chemical) are available for Sand gazelles (see
that chapter), but none have been gathered for the present species.
17)
Other resources
The research department (CRES) of the Zoological Society of San Diego
has had a long interest in the genetics of gazelles, and, especially in
the polymorphism of goitered gazelles (Kingswood et al., 1994); thus,
cell strains of numerous specimens of Persian gazelles and other related
forms are available from CRES
at the Zoological Society of San Diego by contacting Dr. Oliver Ryder
at oryder@ucsd.edu.
There is a very extensive bibliography on all aspects of sand gazelles
and their related forms. This has been gathered by the King Khalid Wildlife
Research Center, Thumammah and can be made available by the National Commission
for Wildlife Conservation and Development, P.O. Box 61681, Riyadh 11575,
Kingdom of Saudi Arabia.
18) Other remarks - What additional Information is needed?
Early stages of implantation need to be observed, and more information
on the length of the umbilical cord is needed.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society
of San Diego. I appreciate also very much the help of the pathologists
at the San Diego Zoo.
References
Carter, S. and Kingswood, S.: Goitered Gazelle. North American Regional
Studbook First edition. Sedgwick County Zoo & Botanical Garden, Wichita,
Kansas, 1991.
Effron, M., Bogart, M.H., Kumamoto, A.T. and Benirschke, K.: Chromosome studies in the mammalian subfamily Antilopinae. Genetica 46:419-444, 1976.
Granjon, L., Vassart, M., Greth, A. and Cribiu, E.-P.: Genetic study of sand gazelles (Gazella subgutturosa marica) from Saudi Arabia. Chromosomal and isozymic data. Z. Säugetierk. 56:169-176, 191.
Gray,
A.P.: Mammalian Hybrids. A Check-list with Bibliography. 2nd edition.
Commonwealth Agricultural Bureaux Farnham Royal, Slough, England, 1972.
Griner, L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego, California, 1983.
Groves, C.P.: An introduction to the gazelles. Chinkara: Bulletin of the Gazelle Research Group. 1(1):4-16, 1985.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 10, Folio 504, 1977.
Kingswood, S.C., Kumamoto, A.T., Sudman, P.D., Fletcher, K.C. and Greenbaum, I.F.: Meiosis in chromosomally heteromorphic goitered gazelle, Gazella subgutturosa (Artiodactyla, Bovidae). Chromosome Research 2:37-46, 1994.
Nowak, R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press, Baltimore, 1999.
Orlov, in Shi, L.: Recent trends in mammalian cytogenetics in China. La Kromosoma II-45:1458-1467, 1987.
Rostron, J.: A multivariate statistical study the skull measurements of five taxa of gazelles. Biol. J. Linn. Soc. 4:1-14, 1972.
Sempere, A.J., Brown, N., Pereladova, O.B., Bahloul, K., Lacroix, A. and Soldatova, N.: Comparative analysis of reproductive cycles in female Persian gazelle (Gazella subgutturosa subgutturosa) (Central Asia) and sand gazelle (Gazella subgutturosa marica) (Arabian Peninsula). Gen. Comp. Endocrinol. 121:57-65, 2001.
Vassart M, Greth A, Durand V and Cribiu EP. Chromosomal polymorphism in sand gazelles (Gazella subgutturosa marica). J Heredity 84:478-481, 1993.
Vassart, M., Granjon, L. and Greth, A.: Genetic study of Gazella gazella: chromosomal and allozymic data. C. R. Acad. Sc. III 318:27-33, 1995.
Walther, F.: Die Gazellen und ihre Verwandten. In, Gzimeks Tierleben. Volume 13. Kindler-Verlag Zürich, 1968.
Wurster, D.H.: Sex-chromosome translocations and karyotypes in bovid tribes. Cytogenetics 11:197-207, 1972.