|
|
(Clicking
on the thumbnail images below will launch a new window and a larger
version of the thumbnail.)
|
Ateles species
Order: Primates
Family: Cebidae
1) General zoological data of species
The term "spider monkey" is used loosely here, as I well appreciate the difficulty in taxonomy. Konstant (1985; 1986) has stated that this designation best, or most properly, applies for Ateles geoffroy,i but that "spider monkey", has now been used for many other, similar-appearing, animals. They are all characterized by their prehensile tail, their body size and black feet. A comprehensive review of these animals was published by Kellogg and Goldman (1944), to be followed by many others. Still, much controversy on taxonomy exists.
This characteristically South American platyrrhine group of species is widely distributed through the Amazon basin and most of Central America. It is composed of many species and/or subspecies that differ greatly in color and some minor other features. Mittermeier (1986) discussed their distribution and status of endangerment in some detail. Despite a lack of publicity, a number of these species/subspecies are critically endangered (Konstant and Mittermeier, 1985). Schneider (2000) compared DNA databases of the major groups of South American primates (Atelidae, Pitheciidae, Cebidae) and further defined their relationships.
Spider monkeys are extremely nimble, prehensile-tailed animals and are often exhibited in zoological parks. They are very social animals. Often large groups travel in the trees of the rainforest. More often, they form smaller family groups. An extensive literature exists on these species that can be accessed through Nowak's book (1999) and the publication by Mittermeier (1986). A clear differentiation among the great array of phenotypes displayed by these animals has been difficult. A crisper designation of phenotypes was attempted by Konstant & Mittermeier (1985; 1986) when they discussed the need for subspecific recognition because of their chromosomal differences and their many different phenotypes. The importance of this was stressed especially for their captive management. They depicted these different phenotypes in color pamphlets (1985) but the work that was meant to come to a unifying agreement was never completed. The same can be said about the many chromosomal forms that are displayed by spider monkeys from different regions. There are many inversions and translocations, which have not found universal acceptance in their correlation with agreed-upon designations. Nor are they used for a more meaningful management (de Boer, 1974). One reason for the difficulty is that the precise origin of many zoo specimens is unknown, and fieldwork with cytogenetic techniques is problematic. More information on recent phenotype/genotype correlation is found below under the section on genetics. For the understanding of placentation all this is not problematic as the reproductive features of all Ateles species is much the same.
2) General gestational data
Adult spider monkeys weigh about 6-7 kg but, occasionally, twice as much.
One of their unusual physical characteristics is the long "clitoris"
of females that makes them appear to be males, especially since the testes
are small and difficult to see. The function of this pendulous clitoris
is unknown, but it is non-erectile. Following mating the male often touches
this organ and smells it. The maximal life span recorded for Ateles
is 33 years.
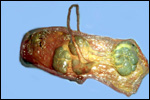
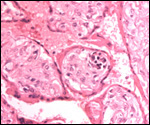
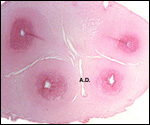
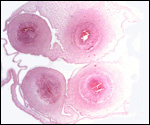
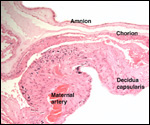
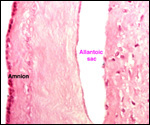
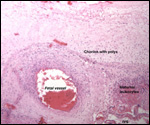
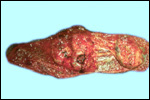
We have had 25 placentas available from term pregnancies, all were bilobed
and the average weight without cord and membranes was 105 g (80-135 g).
In all but one was the primary lobe (the one with cord insertion) the
larger and heavier lobe. It usually weighs 50-75 g and measures 8 cm in
diameter and 1-1.5 cm in thickness; the secondary lobe is only slightly
smaller. One of my placentas had a tiny third lobe. Blood vessels connect
the lobes.
The length of gestation in spider monkeys has been studied in some detail
by Eisenberg (1973). He found it to be 7-7½ months (215-226 days).
The usually single newborn weighs around 340-480 g (Puschmann, 1989).
Sexual maturity is reached between 4-5 years.