 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last updated: March 21, 2006. |
Lissodelphis borealis
Order: Cetacea
Family: Delphinidae
1) General Zoological Data
This species should not be confused with the right whale ( Eubalanea glacialis ), irrespective of their perhaps confusing names; both species (Right Whale & Right Whale Dolphins), however, characteristically lack a dorsal fin. For that reason the right whale dolphins are also called Glattdelphine in German. There are two right whale dolphin species, the one inhabiting the Northern Pacific Ocean is Lissodelphis borealis , and the one of the Southern Hemisphere is L. peronii . Mature females weigh around 80 kg (Nowak, 1999). The North Pacific animals are mostly black and have a white patch of skin on their chest and a white band midventrally. Groups of up to 3,000 have been observed, but they usually travel in groups of 100-200. Newborns measure 60-100 cm in length and are generally found in November. Nishiwaki (1972) provided details of the two species, as well as pictures and dental formulae. Maturity is reached at age 10 years and calving occurs every other year; calves are about ½ the length of adults. They are very social animals. Captive colonies do not exist and virtually nothing substantial is known about their reproductive biology. They feed on fish, are occasionally caught in fishing nets but they are not actively hunted. Their population has dwindled considerably because of the predation by fishing. The Latin name of the right whale dolphin derives from lissos (Gr = smooth), delphis (Gr = dolphin) and boreus (L = northern) according to Gotch (1979).
 |
Appearance of an adult Lissodelphis borealis copied from Slijper & Heinemann in Grzimeks Tierleben. |
2) General Gestational Data
Many reproductive parameters of dolphins are similar and these are summarized in the chapter of the bottle-nosed dolphin that should be consulted. As is common in cetacean, the left uterus and ovary were employed for gestation (Slijper, 1949). Singletons are usually found; I know of no record of twinning in this species.
3) Implantation
A single specimen became available of a pregnant uterus from a stranded dolphin through the courtesy of Dr. Judy St. Leger. It contained a small well-preserved fetus and a mostly detached placenta. The pregnant left uterine horn measured 64 cm in length and 9 cm at its widest diameter. The non-pregnant right horn was 46 x 6 cm. The left ovary contained the active corpus luteum, the right ovary was without corpora albicantia. The entire uterus with cervix weighed 550 g. The female fetus was 11 cm in total length and had a 3 cm long tail. It weighed 23 g and had already small areas of black skin, mainly at the head. Upon radiography, no bony development could be demonstrated; all future bones were still cartilaginous. Initially this fetus was identified as male because of what appeared to be a penile development (see picture below). Histologically, however this is a female fetus with uterus etc. This mistake in gender identification of small fetuses is not unlike what is often found in human fetuses where male gender is assigned to small human female fetuses that normally have a much enlarged clitoris. The placenta was left within the uterus for fixation after removal of the fetus. The allantoic sac with thin amnionic sac cover extended for a short distance into the right uterus.
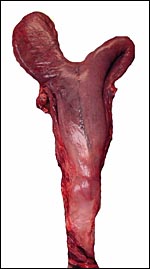 |
Pregnant left uterine horn of right whale dolphin. |
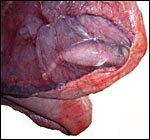 |
Opened uterus with the 11 cm fetus inside its amnionic sac; the latter is overlain by the vasculature of the allantoic sac. Note the positioning of the fetus, tail towards the cervix in a manner of its future birth. |
 |
Female fetus lacking dorsal fin and showing the beginning black coloration at the head. |
 |
Side view of fetus. |
4) General Characterization of the Placenta
This is a diffuse microvillous epitheliochorial placenta with a single layer of trophoblast covering the villi diffusely. It is not invasive.
5) Details of fetal/maternal barrier
The cetacean all have an epitheliochorial placental relationship to the maternal uterus that is exemplified in the next photographs...
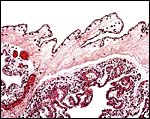 |
Placental surface with amnion above and villi below. |
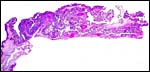 |
Low power survey of villous placenta. |
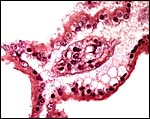 |
Single branching villus with single layer of trophoblast that has microvillous surfaces. |
6) Umbilical cord
The umbilical cord was 7 cm long and had a slight left twist. It contained two large arteries and two veins, numerous tiny vessels and a large, patent allantoic duct. There is only a sparse amount of Wharton's jelly.
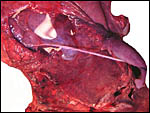 |
The 11 cm fetus (at right) is attached to the placenta by a 9 cm umbilical cord. |
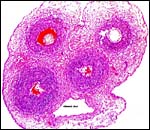 |
Cross section of umbilical cord with large allantoic duct below that is situated between the two arteries. |
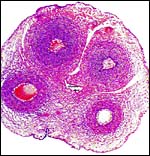 |
In this section the allantoic duct is more centrally located. Tiny vessels are also seen. |
7) Uteroplacental circulation
No studies have been published on this species.
8) Extraplacental membranes
A large, diffusely vascularized allantoic sac surrounds the delicate amnion and it extended into the right uterine horn. The amnionic and allantoic sacs were filled with clear and minimally pale-yellow fluid.
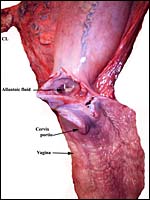 |
This demonstrates the vagina (below), the cervix, endocervix and a partially opened left uterine horn with an arrow pointing at protruding Allantoic sac. |
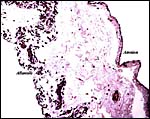 |
Cross section of membranes with allantois at left, amnion at right. |
9) Trophoblast external to barrier
There is no extraplacental trophoblast in endometrium or uterus.
10) Endometrium
The vagina was 14 cm in length and had external black pigmentation where it becomes external skin. The cervical os had a flower-like aperture and the endocervical canal was filled with thick, tenacious mucus. There is no decidua in cetacea.
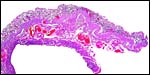 |
Uterine wall with endometrium above. |
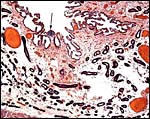 |
Higher magnification of the endometrium with arrow pointing at one of many collections of lymphocytes. The villi would interdigitate with the surface. |
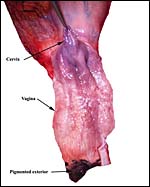 |
The exterior black pigmentation of the exterior vagina can be seen at bottom, the cervical os is indicated by arrow as well. |
11) Various features
There is no decidua in cetacean and neither are there trophoblastic cells.
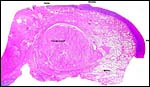 |
This section shows the dermal aperture of the vagina at right and the beginning of the vaginal canal. |
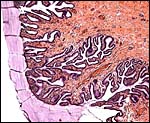 |
The endocervical canal is filled with thick mucus that emanates from the glands. |
12) Endocrinology
As is the commonest finding in cetacean, the left ovary and uterine horns are involved in the reproductive activity (Slijper, 1949). Thus, in this animal also, the left ovary contained the 2x1.2 cm protruding corpus luteum. The right ovary had a smooth surface and no corpora albicantia. As we have suggested previously, the left ovary also contains the corpora albicantia of earlier gestations (Benirschke et al., 1980). Hobson & Wide (1986) have found placental gonadotropins in the placenta of Tursiops that was inactivated with anti-hCG but such studies have not been undertaken in Lissodelphis. The remarkable hypertrophy of the clitoris is as yet unexplained from an endocrinological point of view.
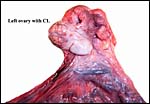 |
The left ovary contains the protruding large corpus luteum. |
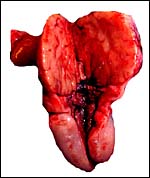 |
The corpus luteum (above) is dark yellow. |
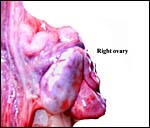 |
The right ovary was inactive although it is filled with primordial ova. |
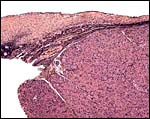 |
This is a cross section of the edge of the corpus luteum (right) and adjacent ovarian cortex. |
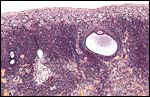 |
There are a few developing follicles in the left ovary and rare corpora atretica (left, center). |
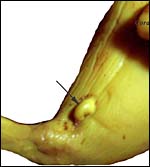 |
Tail portion of fetus with the remarkably prominent clitoris at arrow. (Fixation in Bouin's solution). |
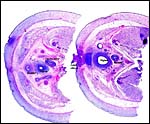 |
Cross sections of pelvis. In the center is the base of the clitoris; vagina and uterus are individually labeled. |
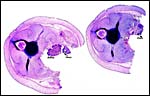 |
Additional cross sections of the abdomen. Ovary and kidney are labeled. |
13) Genetics
Right whale dolphins possess 44 chromosomes, as was first identified by Duffield (1976). Her karyotype was subsequently reproduced by Hsu & Benirschke (1977) and it is here also depicted. Hybrids have not been reported.
 |
Karyotypes from two male Lissodelphis borealis specimens. |
14) Immunology
These features have not been studied. In the cervical mucus are some immunocytes. The vagina has dense collections of immunocytes beneath the epithelium. In addition, focal collections of immunocytes were found in the endometrium. There was no inflammation, however. Minh et al. (2000a) found high concentrations of PCBs in the blubber of many cetaceans, including the right whale dolphin; the TEQ concentration exceeded levels associated with immunosuppression in harbor seals.
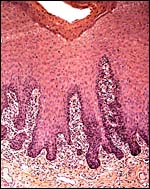 |
Vaginal epithelium with dense collection of immunocytes. |
15) Pathological features
Moser & Rhinehart (1993) identified of a Lissodelphis calf contained worms with characteristics of Halocercus . These authors also studied the potential transplacental transmission of this nematode, but found no such evidence. Woodhouse & Rennie (1991) described vaginal calculi in three dolphin species and studied (by CAT scans) an early gestational right whale dolphin and suggest the possible origin of vaginal calculi from aborted fetuses.
Minh et al. (2000b) found high concentrations of organochlorines in Lissodelphis and attributed it to the widespread region of these cetaceans.
16) Physiologic data
The fetal fluids and blood contain mostly fructose, as is true of Artiodactyla (Slijper, 1972). I am not aware of any other physiological studies. Koopman et al. (2003) evaluated the concentration of isovaleric acid in the blubber (and especially the melon) of a large number of cetaceans; the compound is believed to have importance in echolocation.
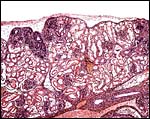 |
Multilobulated fetal kidney. |
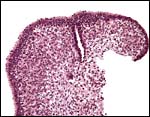 |
Low power appearance of fetal ovary. |
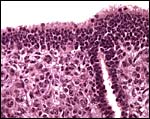 |
Higher magnification of fetal ovary with numerous germ cells. |
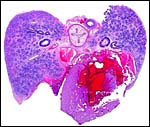 |
Cross section of heart, lung, esophagus and aorta. |
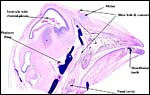 |
Sagittal section of head with individual labels. |
17) Other resources
Cell lines from this fetus were established at CRES and can be made available by contacting CRES at the San Diego Zoo.
18) Other remarks What additional Information is needed?
Mature placentas have not been studied; their weight and size are unknown.
Acknowledgement
The uterus and fetus were kindly made available by Dr. Judy St. Leger of Sea World San Diego, California.
References
Benirschke, K., Johnson, M.L. and Benirschke, R.J.: Is ovulation in dolphins, Stenella longirostris and Stenella attenuata, always copulation-induced? Fishery Bulletin 78:507-528, 1980.
Duffield, D.A.: Phylokaryotypic evaluation of the cetacean. Ph.D. Thesis, University of California Los Angeles , 1976.
Gotch, A.F.: Mammals Their Latin Names Explained. Blandford Press, Poole, Dorset , 1979.
Hobson, B.M. and Wide, L.: Gonadotrophin in the term placenta of the dolphin (Tursiops truncatus), the Californian sea lion (Zalophus californianus), the grey seal (Halichoerus grypus) and man. J. Reprod. Fertil. 76:637-644, 1986.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 10, Folio 489. Springer-Verlag, N.Y. , 1977.
Koopman, H.N., Iverson, S.J. and Read, A.J.: High concentrations of isovaleric acid in the fats of odontocetes: variation and patterns of accumulation in blubber vs. stability in the melon. J. Comp. Physiol. (B) 173:247-261, 2003.
Minh, T.B., Nakata, H., Watanabe, M., Tanabe, S., Miyazaki , N., Jefferson , T.A., Prudente, M. and Subramanian, A.: Isomer-specific accumulation and toxic assessment of polychlorinated biphenyls, including coplanar congeners, in cetaceans from the North Pacific and Asian coastal waters. Arch. Environm. Contam. Toxicol. 39:398-410, 2000a.
Minh, T.B., Watanabe, M., Tanabe, S., Miyazaki , N., Jefferson , T.A., Prudente, M.S., Subramanian, A. and Karuppiah, S.: Widespread contamination by tris(4-chlorophenyl)methane and tris(4-chlorophenyl) methanol in cetaceans from the North Pacific and Asian coastal waters. Environm. Poll. 110:459-468, 2000b.
Moser, M. and Rhinehart, H.: The lungworm, Halocercus sp. (Nematoda: Pseudaliidae) in cetaceans from California . J. Wildl. Dis. 29:507-508, 1993.
Nishiwaki, M.: General Biology. Chapter 1 in Mammals of the Sea. Biology and Medicine. S. Ridgway, ed. Charles C. Thomas, Springfield , Illinois , 1972.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Slijper, E.J.: On some phenomena concerning pregnancy and parturition of the Cetacea. Bijdragen tot de Dierkunde 28:415-448, 1949.
Slijper, E.J.: Functional Morphology of the Reproductive System in Cetacea. Chapter 15, in: Mammals of the Sea. Biology and Medicine. S. Ridgway, ed. Charles C. Thomas, Springfield , Illinois , 1972.
Slijper, E.J. and Heinemann, D.: Die Zahnwale. Chapter 16 in, Grzimeks Tierleben. Vol. 11, Kindler Verlag, Zürich, 1969.
Woodhouse, C.D. and Rennie, C.J 3 rd . Observations of vaginal calculi in dolphins. J. Wildl. Dis. 27:421-427, 1991.