 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last updated: Feb 7, 2008. |
Panthera leo
Order: Carnivora
Family: Pantheridae [Felidae]
1) General Zoological Data
In the past, lions were comprised among the Felidae; more recently, however, a separate genus was assigned, Panthera. Thenius (1967) provided a detailed overview of the skeletal history and general evolutionary aspects of Felidae and he then already clearly separated Panthera from Felis. When O'Brien et al. (1987a) studied electrophoretically the allozymes of pure African and Asian (Gir forest) lions; they found that the Asian lions were monomorphic in 46 loci, while there was moderate variation in the African stock. Nevertheless, the study of the Asian lions showed that two of the original five founders were descendants from an African subspecies, while three others were pure Asian. They made recommendation for the SSP (Species Survival Plan) on this basis. Essentially the same information was made available in a more popular version that contains excellent diagrams (O'Brien, 1987b). Female lions weigh between 100 and 182 kg and the longevity in zoos has been cited as being 13 years; but some lions have lived as long as 30 years (Nowak, 1999).
 |
Male African lion at San Diego Zoo. |
 |
Female African lion at San Diego Zoo's Wild Animal Park . |
2) General Gestational Data
Lions are polyestrous, with heat lasting 4 days; the litters contain one to nine young, usually 3-4. The primigravid female whose placentas are here described had arrested labor with dystocia of the first, larger-than-normal, cub. It was manually extracted but stillborn. The second cub was then delivered by Cesarean section and survived. It was heavily meconium-stained and somewhat smaller that the first.
Gestation in lions lasts 100 to 119 days, and the newborns usually weigh 1,300 g; for this pregnancy, however, it was judged that the pregnancy was 2 weeks overdue, perhaps accounting for the problems of large size and dystocia. Sexual maturity in lions is usually attained at 3-4 years. An additional placenta became available in November 2007. It is the placenta of one of a litter of four normal offspring.
3) Implantation
Numerous publications have described early cat implantation and these were summarized by Denker et al. (1978). No data of lion placentation are available, however. Thus, I assume that the process is generally similar to that of the cat some of which data are here repeated. The previous authors describing cat placentation suggested that, as in other species, there is dissolution (not rupture) of the zona pellucida and those proteinases are responsible for implantation. The blastocyst becomes much expanded before implantation and is oval. The embryonic disk attaches antimesometrially. There is an early yolk sac placentation that is soon replaced by the large allantoic placenta. The allantoic sac fuses with the amnion and this thin allantoic membrane gets to possess blood vessels that are classically lacking from the amnionic membrane. A girdle-like trophoblastic invasion occurs that produces the final placental shape. There is early invasion of the endometrial glandular mouths by trophoblast. This is particularly well illustrated by Kehrer (1973). He also showed the growth of the lamellar tongues of placental tissue in relation to the uterine axis.
Later, the endometrial glands distend with secretion and contain degenerating epithelium. Adjacent to the placental girdle, small hematomas of maternal blood develop. They are, however, much smaller in cats than those found in dog placentas and even less pronounced in lions. The portions of placenta lateral to the girdle are referred to as paraplacenta. At this site, the trophoblast adheres to the uterine epithelium, thereby making this area a truly epithelio-chorial relation. Rosettes, or chorionic vesicles, found here are numerous. Malassiné (1977) studied the erythrocytophagocytosis of maternal red blood cells by trophoblast in cats at this site and suggested that this is an important site of iron absorption. This is, of course, the green/brown border of the zonary placental tissue of carnivores that is so minimal in the lion placenta. Whether or not much other transport occurs across this region is uncertain.
4) General Characterization of the Placenta
Numerous studies have been accorded to cat placentation, but the lion placenta is apparently rarely available. I have been unable to find a written account of its structure. There are many features of this lion placenta, however, that are also seen in the placenta of the tiger (see that species entry). Moreover, the features of placentation in the domestic cats have also many similarities to that of the lion. Thus, the reader should consult with those species to make comparative observations.
The lobulation of this lion placenta is very distinct, as will be see in the photographs below. The male fetus of this placenta (one of twins, the one that was born first) was rather large, weighing 1,875 g. It presented as breech and had to be extracted; the second twin was delivered by Cesarean section and was severely meconium-stained. It weighed 1,620 g and is surviving, albeit not being nursed by the dam. The zonary placenta, including the membranes of the first twin weighed 575 g and measured 42 cm in circumference, 11 cm in width and 0.8 cm in greatest thickness. This is a typical zonary (ring-shaped) placenta with virtually no paraplacental material. The placenta of the second twin, who survived, weighed 425 g and measured 34 x 11 cm. The new placenta weighed 267.5 g (10x36x0.3 cm). It had essentially the same gross and microscopic appearance.
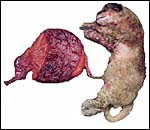 |
Overview of the first specimen available. |
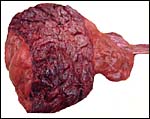 |
The zonary nature of this placenta is evident. |
 |
When the complete circle is cut, this appearance of a multicotyledonary placenta is seen. |
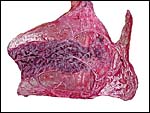 |
Placenta of the second, surviving lion twin. |
5) Details of fetal/maternal barrier
The materno-fetal barrier of cats is endothelial-chorial and the placenta is only superficially invasive into the endometrium but does not extend into the myometrium. A useful description of endotheliochorial placentation has been provided by Dantzer (1999). Enders et al. (1998) further defined the interhemal distance in a variety of placentas. For Felidae they assigned the following characteristics: syncytial trophoblast is adjacent to maternal endothelium'. Mossman (1987) also described the felid placenta as being endotheliochorial, but Zhemkova (1962) took issue with this and declared that the cat placenta is hemochorial. Electron microscopic studies, however, did not confirm Zhemkova's opinion. In comparison with the domestic cat, the tiger and lion placentas have a much denser and more complex structure and the lamellae of the labyrinth of the lion placenta are unusually straight (Mossman, 1987). The lion placental structure thus exhibits very much similarity to that of the tiger's.
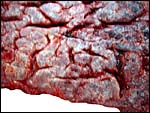 |
The maternal surface has numerous clefts that correspond to individual cotyledons that can be seen from above. |
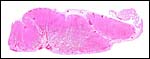 |
Cross-section of three adjacent lobules in lion placenta. Separation from uterus is below. |
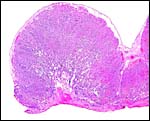 |
Another lion placental lobule in cross section. Implantation site below. |
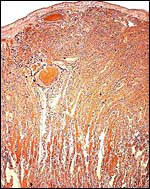 |
Upper portion of the labyrinth with nearly parallel, straight lamellae. Note the one large maternal blood channel. |
 |
The labyrinth becomes more loosely structured near the placental floor; the lamellae are more separated by large maternal vascular channels. |
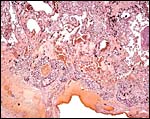 |
Placental floor. Note trophoblast giant cells surrounding maternal vessels. |
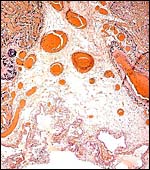 |
Floor of lion placenta in the center of a microcotyledon. Large maternal blood vessels of endometrium; villous tissue at top. |
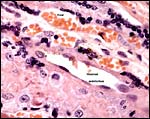 |
Maternal and fetal blood is separated by a small amount of connective tissue and endothelia. |
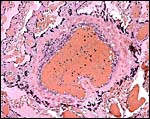 |
Trophoblastic giant cells surround a major maternal vessel as it delivers blood to the internal portion of the disk. |
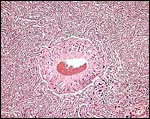 |
Large maternal vessel surrounded by trophoblastic giant cells; center of labyrinth. |
6) Umbilical cord
The umbilical cord of this specimen was marginally attached, measured 17 cm and contained four vessels (2 arteries and 2 veins) plus a widely patent allantoic duct. There were no spirals and no foci of squamous metaplasia were present. The two veins course in the abdomen, one on the right to the liver hilus, the other to the left vena cava. The arteries followed their usual course next to the bladder. As will be seen below, as the vessels divided over the fetal surface they were contained in folded membranes.
 |
The 17 cm long cord is not twisted and approximately one half is amnionic, the other allantoic. |
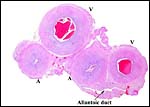 |
Cross section of umbilical cord. |
 |
The cord divides its main blood vessels within the membranes. |
7) Uteroplacental circulation
No relevant information is available other than that which can be gleaned from the histologic analysis of the placenta.
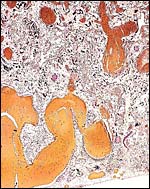 |
Hugely distended maternal vessels at the site of separation of lion placenta. From these, large channels deliver blood into the internal labyrinth. |
8) Extraplacental membranes
The amnion develops by folding. Its epithelium is very thin and has no vasculature. It is attached to the chorion and, in some areas, to the large allantoic sac. The allantoic membrane has an extensive vasculature and a relatively thick epithelium that becomes thinner with advancing gestation. In this placenta, the membranes were filled with fluid (contained in the allantoic sac) but the fetal stomach was also massively distended with ingested amnionic fluid. There is a minimally brown pigment attached to the maternal surface of the membranes, presumably corresponding to material of the hemophagous region' of other felids (Malassiné (1977). The deep green pigment, as seen in dogs and less so in domestic cats, is absent. When Prussian blue stains were done, only a minimal amount of blue staining resulted, but no typical hemosiderin granules were found. The details of iron recovery by the fetus from maternal hemorrhages at the lateral aspect of the feline placenta have been sketched in excellent detail and with line drawings in a contribution by Burton (1982). While minute amount of the pigment are present at the lion girdle's margin, it is not impressive.
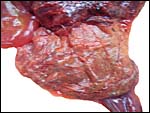 |
The external aspect of the membranes had a rust-colored granular aspect. |
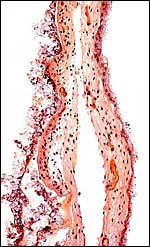 |
Membranes near the margin of the girdle; at left, some maternal endometrial endothelial cells are attached. |
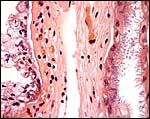 |
Higher magnification of membranes' with remnants of maternal endometrium and trophoblast at left, allantois at right. |
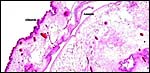 |
The allantoic membrane is heavily vascularized, not the amnion. |
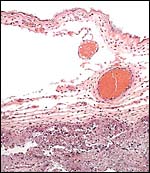 |
Membranes adjacent to placental girdle showing much endometrial debris (some pigment) at lower right bottom. |
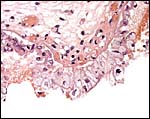 |
Higher magnification. Trophoblast and endometrial debris at edge of girdle. |
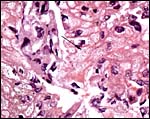 |
Minute granular content in cytoplasm of trophoblast and endometrium. |
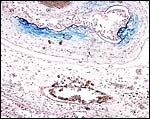 |
Iron stains showing some diffuse minor deposits of Prussian blue staining at margin of placenta. |
9) Trophoblast external to barrier
Since nidation is quite superficial, there are no invasive trophoblastic cells in the myometrium.
10) Endometrium
Felidae have a bicornuate uterus with a short body but long horns. Decidua does not develop in them.
11) Various features
No special features are known to me.
12) Endocrinology
Only one of the testes of the first tiger dissected here was descended despite its size and possible postmaturity, the other lay in the inguinal canal. The ovaries of lions are contained in a bursa and all felids have essentially similar ovaries (Mossman & Duke, 1973); and females ovulate upon copulation.
13) Genetics
Lions have 38 chromosomes (Wallace & Fairall, 1967). Roubin et al. (1973) compared banding patterns of various felidae and established that tiger and lion had a pericentric inversion of chromosome #6. Wurster-Hill & Gray (1973) also described the banding pattern and compared it with other felidae. Tian et al. (2004) showed by chromosome painting study that the lion and clouded leopard have identical karyotypes. Satoh et al. (1982) ascribed the nucleolus-organizing region to one small submetacentric element (E 1 of the San Juan convention on cat chromosomes). Moore (1965) reviewed the sexual dimorphism of lion cells, the technique by which the original Barr-body was discovered, which he reviewed also.
Collier & O'Brien (1987) considered the phylogeny of felidae by investigating their immunological distances. More recent information on genes and linkage maps are contained in a report of a symposium on Advances in Canine and Feline Genomics: Comparative Genome Anatomy and Genetic Disease in J. Hered. 94, Issue 1 (January), 2003, but the majority of this was dedicated to canine species. Hybridization with tigers occurs (Gray, 1972) and hybrids with leopards have also been reported to occur in zoos. Rife (1965) is more skeptical of such hybrids. Generally tiglons' and ligers' are fertile only in the female hybrids (Rife (1965). A mounted specimen of an alleged hybrid with a jaguar was discussed by Gray.
 |
Karyotype of male African lion (CRES). |
14) Immunology
The immunologic distance of felidae was described in detail by Collier & O'Brien (1985) that separated tiger and lion from other felidae. I am not aware of truly immunological studies undertaken in lions.
15) Pathological features
Griner (1983) autopsied a large number of lions and found that we had numerous neonatal deaths or stillbirths. He also grew fungi from a neonatally developing skin condition that matted hair and became scaly. Others had aspiration pneumonia, and one litter of four had cleft palates in all offspring. Heuschele (1959) reported on another cub with cleft palate. Griner also saw a hepatic lymphosarcoma in a 15 year-old female, but he reported no parasites. Stroud & Coles (1980) described death from blastomycosis in an African lion that had been transferred to Oregon from Tennessee . It had widespread lesions but cultures were not successful because of freezing the tissues.
Brückner & Lang (1948) described in detail the removal of a cataract from a lioness, including the histologic findings of the lens. O'Sullivan et al. (1977) described neurologic lesions secondary to thick skull bones that led to compression of vermis and cerebellum with herniation and medulla, in young captive lions that were presumed to be due to Vitamin A deficiency.
16) Physiologic data
Wittermeyer Mills (1980) studied the effect of two different diets (frozen meat and dry diet) on lions and tigers. She found that the dry diet led to diarrhea, although it was less expensive. Lang & Bachmann (1956) summarized their results of a Cesarean section on a triplet gestation in which the fetuses were estimated to have died 2-3 days earlier. Earlier, Lang (1945) had described major details of anesthesia in lions and tigers.
17) Other resources
Cell lines are available from the Frozen Zoo of CRES at the Zoological Society of San Diego.
18) Other remarks
Since I was unable to find any publications on lion placentas, and especially none on early implantation, it would be desirable if these were described in the future.
Acknowledgement
The animal photographs in this chapter and the karyotype come from the Zoological Society of San Diego.
References
Brückner, R. and Lang, E.M.: Kataraktextraktion bei einer Löwin. Schweiz. Arch. Tierheilk. 90:582-594, 1948.
Burton , G.J.: Review article. Placental uptake of maternal erythrocytes: a comparative study. Placenta 3:407-433, 1982.
Collier, G.E. and O'Brien, S.J.: A molecular phylogeny of the felidae: Immunological distance. Evolution 39:473-487, 1987.
Dantzer, V.: Endotheliochorial placentation. Pp.1078-1084, in Encyclopedia of Reproduction. Knobil and J.D. Neill, eds. Vol. 1. Academic Press, San Diego , CA , 1999.
Denker, H.-W., Eng, L.A. and Hammer, C.E.: Studies on the early development and implantation in the cat. II. Implantation: Proteinases. Anat. Embryol. 154:39-54, 1978.
Enders, A.C., Blankenship, T.N., Lantz, K.C. and Enders, S.S.: Morphological variation in the interhemal areas of chorioallantoic placentae. A review. Trophoblast Res. 12:1-19, 1998.
Gray, A. P.: Mammalian Hybrids. A Check-list with Bibliography. 2nd ed. Commonwealth Agricultural Bureaux, Farnham Royal, 1972.
Griner, L.A. : Pathology of Zoo Animals. Zoological Society of San Diego , San Diego , California , 1983.
Heuschele, W.P.: Cleft palate in lions of one litter A case report. J. Amer. Vet. Assoc. 134:365-366, 1959.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Springer-Verlag , New York .
Kehrer, A.: Zur Entwicklung und Ausbildung des Chorions der Placenta zonaria bei Katze, Hund und Fuchs. Z. Anat. Entwickl.-Gesch. 143:25-42, 1973.
Lang, E.M.: Die Narkose bei Löwe und Tieger. Schweiz. Arch. Tierheilk. 87:486-496, 1945.
Lang, E.M. and Bachmann, H.: Kaiserschnitt bei einer Löwin. Schweiz. Arch. Tierheilk. 98:111-115, 1956.
Malassiné, A.: Étude ultrastructurale du paraplacenta de chatte: mécanisme de l'érythrocytophagocytose par la cellule chorionique. Anat. Embryol. 151:267-283, 1977.
Moore , K.L.: Sexual dimorphism in interphase nuclei of the lion, Felis leo. Canad. J. Zool. 43:439-445, 1965.
Mossman, H.W.: Vertebrate Fetal Membranes. MacMillan, Houndmills, 1987.
Mossman, H.W. and Duke, K.L.: Comparative Morphology of the Mammalian Ovary. University of Wisconsin Press, Madison , Wisconsin , 1973.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
O'Brien, S.J., Joslin, P., Smith III, G.L.., Wolfe, R., Schaffer, N., Heath, E., Ott-Joslin, J., Rawal, P.P., Bhattacharjee, K.K. and Martenson, J.S.: Evidence for African origins of founders of the Asiatic lion species survival plan. Zoo Biol. 6:99-116, 1987a.
O'Brien, S.J., Martenson, J.S., Packer, C., Herbst, L., de Vos, V., Joslin, P., Ott-Joslin, J., Wildt, D.E. and Bush, M.: Biochemical genetic variations in geographic isolates of African and Asiatic lions. Natnl. Geograph. Res. 3:114-124, 1987b.
O'Sullivan, B.M., Mayo, F.D. and Hartley, W.J.: Neurologic lesions in young captive lions associated with vitamin A deficiency. Austral. Vet. J. 53:187-189, 1977.
Rife, D.C.: Hybrids. Public Affairs Press, Washington , D.C. 1965.
Roubin, M., de Grouchy, J. and Klein, M.: Les félidés: Évolution chromosomique. Ann. Génét. 16:233-245, 1973.
Satoh, H., Tsuchiya, K. and Hori, H.: The location of nucleolus organizing regions in somatic chromosomes of the African lion, Panthera leo . Chromosome Inform. Serv. 32:28-29, 1982).
Stroud, R.K. and Coles, B.M.: Blastomycosis in an African lion. J. Amer. Vet. Assoc. 177:842-844, 1980.
Thenius, E.: Zur Phylogenie der Feliden (Carnivora, Mamm.). Z. Zool. Syst. Evolutionsforschung 5:129-143, 1967.
Tian, Y., Nie, W., Wang, J., Ferguson-Smith, M.A. and Yang, F.: Chromosome evolution in bears: reconstructing phylogenetic relationships by cross-species chromosome painting. Chromos. Res. 12:55-63, 2004.
Wallace, C. and Fairall, N.: Chromosome studies in an African lion. The Leech 37:16-18, 1967.
Wittmeyer Mills, A.: A comparative study of the digestibility and economy of three feline diets when fed to lions and tigers in confinement. Pp. 87-91 in, The Comparative Pathology of Zoo Animals. R.J. Montali and G. Migaki eds., Smithsonian Institution Press, Washington D.C. , 1980.
Wurster-Hill, D.H. and Gray, C.W.: Giemsa banding patterns in the chromosomes of twelve species of cats (Felidae). Cytogenet. Cell Genet. 12:377-397, 1973.
Zhemkova, Z.P.: The use of sex chromatin in identifying embryonic and maternal tissues in the placenta: New observations on the haemochorial nature of the cat placenta. J. Embryol. Exp. Med. 10:127-139, 1962.