 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last updated: Sept. 21, 2011. |
Orcinus orca
Order: Cetacea
Family: Delphinidae
1) General Zoological Data
There is a single species of Orcinus that populates all oceans; the former subdivisions have been abandoned. This is the largest member of the Delphinidae. In addition to their wide distribution, numerous aquariums exhibit killer whales and many breed this species. This social animal occurs in schools of up to 250 animals and it is also easily trained' to perform. Females weigh up to 5,500 kg, males up to 9,000 kg, according to Nowak (1999). Weigl (2006) described maximum longevity as at least 34 years, but Nowak indicated longevity to 50-60 years. Gotch (1979) derived the name from the Latin: whale=orca .
Thewissen ed. (1998) suggested that the earliest whales specimens recovered are from 50 MYA and he traced the development of these animals from the time of their appearance in a remarkable book. Recent studies by Nikaido et al. (1999) have shown the close genetic relationship between hippopotamus and cetacea. A wide variety of web sites and internet references exist for this species.
 |
Killer whale with calf. |
2) General Gestational Data
The single newborns weigh around 180 kg and are born after a gestation period of 517 days (Nowak, 1999). Robeck et al. (2004) researched the reproductive and endocrine physiology of killer whales with the intent of performing artificial insemination. They did this twice, and they also described the semen quality.
3) Implantation
The cetacean placenta is an epitheliochorial, diffuse organ that implants all over both uterine horns. It fills the entire uterus, with the umbilical cord inserting near the endocervical os. The time of implantation following fertilization, however, remains unknown. The main body of the fetus occupies one horn, while the tail occasionally extends into the other horn. The intrauterine position of the cetacean fetus has been illustrated by Slijper (1956, 1966). The position of the fetus is generally such that the tail delivers first. Wislocki & Enders (1941) described the placenta of bottlenose dolphins (see the chapter on dolphins).
4) General Characterization of the Placenta
I have had three specimens of killer whale placentas for examination (Benirschke & Cornell, 1987), the only reports other than that by Turner (1872) who had an entire uterus with fetus for study. In contrast to his remarks about findings naked' ends of the chorionic sac, ours had villi covering these tips. Turner's fetus was located in the left uterine horn, while the corpus luteum was found in the right ovary. The placenta extended into both horn; being smaller in the horn that did not contain the fetus. In general, however, odontocetes have their ovulation in the left ovary (Benirschke et al., 1980).
| Fetal Weight | Placental weight | Cord length | Outcome | |
| 1 | 160 kg | 1,700g | 80 cm | lived |
| 2 | 158 kg (at death) | 2,650 g | died age 11 days | |
| 3 | 155 kg | ? | stillborn | |
 |
Layout of killer whale placenta with central cord insertion and allantoic sac. |
 |
Diagram of this placenta with its measurements (from Benirschke & Cornell, 1987). |
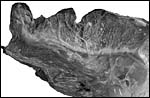 |
The tip' of the placenta (top left) is covered with villi; a distinct ridge extended over the chorionic plate. |
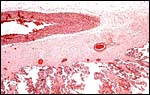 |
Chorion with large vessels and villi extending peripherally. |
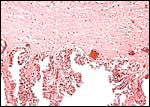 |
Chorion with villi and small chorionic vessels. |
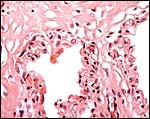 |
Two villi arising from the chorion. They are covered by a single-layered trophoblast layer. |
5) Details of fetal/maternal barrier
The entire villous periphery is covered with a single layer of cuboidal trophoblast; there are no denuded areas. The endometrium is covered with epithelium that is not eroded. Thus, the barrier is complete epitheliochorial. The trophoblast is underlain by numerous fetal capillaries, has fine PAS positive pigment granules, and has frequent vacuoles.
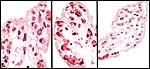 |
Details of villi with single layer of trophoblast and large number of fetal capillaries. |
6) Umbilical cord
The umbilical cord was short, ~80 cm long and had no turns; it was 4 cm in diameter. The cord inserted near the endocervical canal; it apparently ruptured spontaneously during delivery near its insertion on the fetus. The normal umbilical cord contains two large arteries and two large veins, without major anastomoses between any of the vessels. The umbilical vessels branch at the placental insertion dichotomously towards the periphery of the placenta. In addition to the major umbilical vessels, there is a profusion of small and medium-sized blood vessels in the umbilical cord, as many as up ~hundreds in cross-sections. Many have no musculature and are then scattered irregularly through the cord. They have no particular predilection to locate around the allantoic duct. Injection with radio-opaque dyes of the large veins and arteries (undertaken in a bottlenose dolphin) discloses that the small vessels originate from the large ones and also, that they communicate among the large vessels. The function of these small vessels is unknown. The central portion of the umbilical cord carries an irregularly shaped allantoic duct with sparse, and usually very thin epithelium. The Wharton's jelly is sparse and unusually firm. On the umbilical surface, foci of squamous metaplasia (caruncles) are common. They are often remarkably pigmented, both black and white. These plaques' of squamous metaplasia extended some distance onto the amnion but were there more widely disseminated.
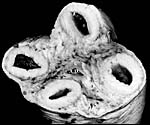 |
Cross section of the umbilical cord with 2 arteries, 2 veins and a central allantoic duct (A.D.). |
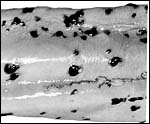 |
Pigmented and unpigmented foci of squamous metaplasia on the surface of the umbilical cord. |
 |
This is the edge of the cord's allantoic duct with low (detached) epithelium. |
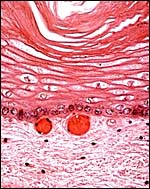 |
Plaque on the cord surface with extensive squamous metaplasia. Note the pigmented basal epithelium and numerous basal capillaries. |
7) Uteroplacental circulation
There are no specific publications dealing with either the uterine circulation or its physiology, but the arrangement of the pelvic vasculature was discussed by Rommel and colleagues (1993). They described that the uterus is surrounded by a complex countercurrent system of blood vessels, which, they suggested, is being used for cooling purposes. There is also an extensive consideration of various other circulatory aspects in their description.
8) Extraplacental membranes
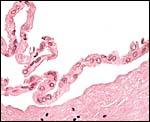
There are no free membranes, but amnion and allantoic sacs are contained within the chorionic sac. The allantoic duct within the umbilical cord is lined by columnar or cuboidal epithelium and is also accompanied by bundles of smooth muscle. This jelly roll shown next was prepared from the interface of the membranes; it has the amnion inside which has a very simple, flat epithelium. There were no vessels in the allantoic membrane. The allantois was cannulated through the allantoic duct in the umbilical cord, thus confirming the arrangement of the diagram shown earlier. Several very small pieces of presumed hippomanes', crumbling material of darker color, were present in the allantoic sac of one specimen.
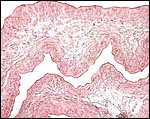 |
Amnionic and allantoic membranes without vessels and composed of connective tissue and thin epithelium. |
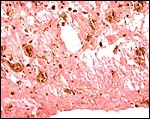 |
Heavily pigmented allantoic debris (hippomanes') of one specimen. |
9) Trophoblast external to barrier
There is no invasion of the uterus by trophoblast.
10) Endometrium
No decidua is present in cetacea. There is an enormous development of superficial capillaries as well as large numbers of racemose glands in the endometrium (Slijper, 1966). The microscopic anatomy of the female genital tract, including the endometrium, has been described for marine mammals in general by Simpson and Gardner (1972). It includes a description of the arterial supply, as well as of the cervix and the ovaries.
11) Various features
The myoglobin of killer whales was studied by Castillo et al. (1977); it resembled that of the sperm whale more than porpoises. Electronmicroscopic studies on semen were performed by Miller et al. (2002).
12) Endocrinology
I am aware of only few endocrine studies in killer whales; however, Hobson & Wide (1986) extracted chorionic gonadotropin from a bottlenose dolphin placenta. Whether delphinidae produce functional placental gonadotropins is currently being examined in more details by Japanese scientists. Robeck et al. (2004) measured estrogens, progestins and LH in killer whales in anticipation of performing artificial insemination. Robeck & Monfort (2006) characterized the male killer whale endocrine physiology with testosterone measurements. More recently, Kusuda et al. (2011) have determined the length of the ovarian cycle in a single killer whale by observing rectal temperature and progesterone levels. By these observations the length of the cycle is approximately 44 days.
13) Genetics
A number of chromosome studies have been performed on killer whales; they are summarized by Hsu & Benirschke (1973) from which the following figure comes. Killer whales have 44 chromosomes; an excellent banding study was done on six killer whales that showed considerable polymorphism of C-bands; this is shown in the following picture (Arnason et al., 1980). Hybrids have not been reported to the best of my knowledge. It is interesting, however, that now a web-based DNA-identification system has been developed (Ross et al., 2003) for ready identification of individual specimens. It is universally available. There have also been studies to ascertain how killer whales in pods avoid inbreeding. On the other hand, Hoelzel et al. (1998; 2002) studied the genetic variation worldwide of killer whales and found relatively little genetic variety.
 |
Karyotypes of male and female specimens (from Hsu & Benirschke, 1973). |
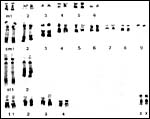 |
Karyotype (C-banded) of female killer whale (from Arnason et al., 1980). |
14) Immunology
I am not aware that any studies have been published other than the review of immunoglobulin concentrations by Taylor et al. (2002).
15) Pathological features
Ridgway (1979) has summarized major causes of death in killer whales from oceanaria and presents a wide variety of lesions. Various infections have been identified in delphinidae, including the killer whale (e.g. Robeck & Dalton, 2002). Brucella infection has been reported from bottlenose dolphins (Miller et al., 1999) and may also occur in killer whales, although direct confirmation from antibody studies is not confirmatory (Omata et al., 2006). Various investigators have identified environmental toxins/contaminants I odontocetes (Minh et al., 2000 a,b; Koopman et al., 2003. In reviewing material published in web sites of cetacea it appears that killer whales often have tapeworms, roundworms and flukes. Also, Hodgkin's disease has been reported (Yonezawa et al., 1989) and severe coronary arteriosclerosis was found in a stranded specimen. Murata et al. (2004) identified toxoplasma antibodies in some captive animals.
16) Physiologic data
There are extensive data collected in the book by Ridgway (1972) for many physiological characters.
17) Other resources
No specific other anatomic studies are reported.
18) Other remarks What additional Information is needed?
Modern studies of an implanted placenta are desirable.
Acknowledgement
The animal photograph in this chapter comes from Dr. Sam Ridgway, San Diego .
References
Arnason, U., Lutley, R. and Sandholt, B.: Banding studies on six killer whales: An account of C-band polymorphism and G-band patterns. Cytogenet. Cell Genet. 28:71-78, 1980.
Benirschke, K., Johnson, M.L. and Benirschke, R.J.: Is ovulation in dolphins, Stenella longirostris and Stenella attenuate, always copulation-induced? Fishery Bulletin 78:507-528, 1980.
Benirschke, K. and Cornell, L. H.: The placenta of the killer whale, Orcinus orca. Marine Mammal Sci. 3:82-86, 1987.
Castillo, O., Lehmann, H. and Jones, L.T.: The myoglobin of the killer whale (Orcinus orca). Biochem. Biophys. Acta 491:23-28, 1977.
Gotch, A.F.: Mammals Their Latin Names Explained. Blandford Press, Poole, Dorset , 1979.
Hobson, B.M. and Wide, L.: Gonadotrophin in the term placenta of the dolphin (Tursiops truncatus), the Californian sea lion (Zalophus californianus), the grey seal (Halichoerus grypus) and man. J. Reprod. Fertil. 76:637-644, 1986.
Hoelzel, A.R., Dahlheim, M. And Stern, S.J.: Low genetic variation among killer whales (Orcinus orca) in the eastern north Pacific and genetic differentiation between foraging specialists. J. Hered. 89:121-128, 1998.
Hoelzel, A.R., Natoli, A., Dahlheim , M.E. , Olavarria, C., Baird, R.W. and Black, N.A.: Low worldwide genetic diversity in the killer whale (Orcinus orca): implications for demographic history. Proc. Biol. Sci. 269:1467-1473, 2002.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 7; Folio 330, 1973.
Koopman, H.N. Iverson, S.J. and Read, A.J.: High concentrations of isovaleric acid in the fats of odontocetes: variation and patterns of accumulation in blubber vs. stability in the melon. J. Comp. Physiol. (B). 173:247-261, 2003.
Kusuda, S., Kakizoe, Y., Kanda, K., Sengoku, T., Fukumoto, Y., Adachi, I., Watanabe, Y. and Doi, O.: Ovarian cycle approach by rectal temperature and fecal progesterone in a female killer whale, Orcinus orca. Zoo Biology 30:285-295, 2011.
Miller, W.G., Adams , L.G., Ficht, T.A., Cheville, N.F., Payeur, J.P., Harley, D.R., House, C. and Ridgway, S.H.: Brucella-induced abortion and infection in bottlenose dolphins (Tursiops truncatus). J. Zoo and Wildlife Med. 30:100-110, 1999.
Miller, D.L., Styer, E.L., Decker, S.J. and Robeck, T.: Ultrastructure of the spermatozoa from three odontocetes: a killer whale (Orcinus orca), a Pacific white-sided dolphin (Lagenorhynchus obliquidens) and beluga (Delphinapterus leucas). Anat. Histol. Embryol. 31:158-168, 2002.
Minh, T.B., Watanabe, M., Tanabe, S., Miyazaki , N., Jefferson , T.A., Prudente, M.S., Subramanian, A. and Karuppiah, S.: Widespread contamination by tris(4-chlorophenyl)methane and tris(4-chlorophenyl)methanol in cetaceans from the North Pacific and Asian coastal water. Environm. Pollut. 110:459-468, 2000a.
Minh, T.B., Nakata, H., Watanabe, M., Tanabe, S., Miyazaki , N., Jefferson , T.A., Prudente, M.S. and Subramanian, A.: Isomer-specific accumulation and toxic assessment of polychlorinated biphenyls, including coplanar congeners, in cetaceans from the North Pacific and Asian coastal waters. Arch. Environm. Contam. Toxicol. 39:398-410, 2000b.
Murata, K., Mizuta, K., Imazu, K., Terasawa, F., Taki, M. and Endoh, T.: The prevalence of Toxoplasma gondii antibodies in wild and captive cetaceans from Japan . J. Parasitol. 90:896-898, 2004.
Nikaido, M., Rooney, A.P. and Okada, N.: Phylogenetic relationships among certartiodactyls based on insertions of short and long interspersed elements: Hippopotamuses are the closest extant relatives of whales. Proc. Natl. Acad. Sci. USA 96:10261-10266, 1999.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Omata, Y., Umeshita, Y., Watarai, M., Tachibana, M., Sasaki, M., Murata, K. and Yamada, T.K.: Investigation for presence of Neospora caninum, Toxoplasma gondii and Brucella -species infection in killer whales (Orcinus orca) mass-stranded on the coast of Shiretoko, Hokkaido, Japan. J. Vet. Med. Sci. 68:523-526, 2006.
Ridgway, S.H.: Reported causes of death of captive killer whales (Orcinus orca). J. Wildl. Dis. 15:99-104, 1979.
Ridgway, S.H., ed.: Mammals of the Sea. Charles C. Thomas, Publisher, Springfield IL , 1972.
Robeck, T.R. and Dalton , L.M.: Saksenaea vasiformis and Apophysomyces elegans zygomatic infections in bottlenose dolphins (Tursiops truncatus), a killer whale (Orcinus orca), and pacific white-sided dolphins ( Lagenorhynchus obliquidens ). J. Zoo Wild. Med. 33:356-366, 2002.
Robeck, T.R. and Monfort, S.L.: Characterization of male killer whale (Orcinus orca) sexual maturation and reproductive seasonality. Theriogenology 66:242-250, 2006.
Robeck, T.R., Steinman, K.J., Gearhart, S., Reidarson, T.R., McBain, J.F. and Monfort, S.L.: Reproductive physiology and development of artificial insemination technology in killer whales (Orcinus orca). Biol. Reprod. 71:650-660, 2004.
Rommel , S.A. , Pabst, D.A. and McLellan, W.A.: Functional morphology of the vascular plexuses associated with the cetacean uterus. Anat. Rec. 237:538-546, 1993.
Ross, H.A., Lento, G.M., Dalebout, M.L., Goode, M., Ewing , G., McLaren, P., Rodrigo, A.G., Lavery, S. and Baker, C.S.: DNA surveillance: Web-based molecular identification of whales, dolphins, and porpoises. J. Hered. 94:111-114, 2003.
Simpson, J.G. and Gardner , M.B.: Comparative microscopic anatomy of selected marine mammals. Chapter 5 in, Mammals of the Sea. Biology and Medicine. S.H. Ridgway, ed. C.C. Thomas, Springfield , Illinois , pp. 298-418, 1972.
Slijper, E.J.: Some remarks on gestation and birth in cetacea and other aquatic mammals. Hvalradets Skrifter 41:1-62, 1956.
Slijper, E.J.: Functional morphology of the reproductive system in Cetacea. Chapter 15 in, Whales, Dolphins, and Porpoises. K.S. Norris, ed. Univ. Calif. Press, Berkeley, 1966.
Taylor , B.C., Brotheridge, R.M., Jessup, D.A. and Stott, J.L.: Measurement of serum immunoglobulin concentration in killer whales and sea otters by radial immunodiffusion. Vet. Immunol. Immunopathol. 89:187-195, 2002.
Thewissen, J.G.M., ed. The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea. Plenum Press, 1998.
Turner, W.: On the gravid uterus and on the arrangement of the foetal membranes in the Cetacea. Transact. Royal Society of Edinburgh, 26:467-504, 1982.
Weigl, R.: Longevity of Mammals in Captivity; From the Living Collections of the World. [Kleine Senckenberg-Reihe 48]. E. Schweizerbart'sche Verlagsbuchhandlung (Nägele und Obermiller), Stuttgart, 2005.
Yonezawa, M., Nakamine, H., Tanaka, T. And Miyaji, T.: Hodgkin's disease in a killer whale (Orcinus orca). J. Comp. Pathol. 100:203-207, 1989.