 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
Presbytis (Semnopithecus) entellus
Order: Primates
Family: Cercopithecidae
1) General Zoological Data
There are 15 species of Presbytis, according to Nowak (1999), all distributed over southern Asia. Hanuman langurs form large groups in India and in several adjacent countries. They are mostly folivorous and inhabit trees but they also walk often on the ground as well. The designation Semnopithecus is preferred by some, but Presbytis is perhaps even more frequently used as well. Nowak (1999) stated that Hanuman langurs are now endangered because of an encroaching human expansion. Many zoos have kept this species, at least for some time, but their maintenance has its challenges and the colonies have often died out. Often this can be attributed to their folivory and inadequate food supply. Gotch (1979) stated that "Hanuman" is a Hindu word for a monkey god. Presbytis refers to an old man, while entellus comes from a figure in Virgil's Aeneid.
 |
Family group of Hanuman langurs at San Diego Zoo's Wild Animal Park. Note the extraordinary length of the tails. |
2) General Gestational
Data
The estrous cycle is 24 - 30 days and estrus lasts 5-6 days. Births occur
at any time during the year. Gestation is from 190-210 days (Roonwal &
Mohnot, 1977). Singleton births are the rule, but twins have also been observed.
The weight at birth is around 500 g, and longevity may be beyond 25 years
(Nowak, 1999).
3) Implantation
I am not aware of any studies of early implantations in Hanuman langurs.
Those for Douc langurs and, especially for the rhesus monkey, can be found
under the relevant chapters. Burton (1980), on the other hand, had the
opportunity of studying two Presbytis obscura (Dusky Leaf Monkeys)
uteri in early gestations and also referred to an old publication by Selenka.
Placentation in that species is likely to be similar to the Hanuman langur
gestation, thus these may be guide posts. The large size of the blastocyst
and its superficial nature of implantation were highlights of the observations
by Burton (1980). He also remarked on the then still absent decidualization
and highlighted the proliferation of endometrium lateral to the invasion
of trophoblast. Cytotrophoblast is also described as infiltrating superficial
maternal blood vessels.
4) General Characterization of the Placenta
I have had over 20 Hanuman langur placentas available for study; some
came from term gestations, a few were from spontaneous abortions. Some
placentas were bilobed, a few had only a single lobe. All placentas had
some old infarcted lesions. Normally, I expect that this animal would
have a bilobed placenta, with the umbilical cord inserting on the larger
lobe. The absence of a second lobe also occurs in about 15% of rhesus
monkeys (see that chapter) and usually does not threaten the gestation.
The average lobe of the placenta measures 8 -12 cm in diameter at term
and is between 0.5 and 1 cm in thickness. The combined weight of the lobes,
without cords and membranes, is given in the table that follows (see also
Benirschke, 1982).
This is a villous placenta with hemochorial relation to the maternal system.
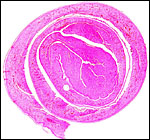
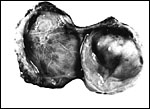 |
Bilobed immature placenta of Hanuman langur, with cord insertion on the larger left lobe. |
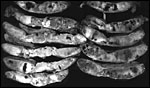 |
Cross sections of mature Hanuman langur placenta showing the large number of infarcts, not unusual and not leading to fetal demise. |
| Placenta
# |
#
of lobes |
Total
weight |
Cord
length |
#
vessels |
Outcome |
| 4370 |
Two |
125
g |
35
cm |
3 |
|
| 4620 |
Two |
140
g |
|
3 |
Infarcts |
| 3564 |
Two |
190
g |
25
cm |
3 |
625 g, infarcts |
| 3565 |
One |
130
g |
25
cm |
2 |
750 g, infarcts |
| 3085 |
Two |
156
g |
30
cm |
3 |
|
| 4908 |
Two |
153
g |
27
cm |
3 |
Infarcts |
| 4899 |
Two |
170
g |
39
cm |
3 |
Infarct |
| 4250 |
Two |
185
g |
44.5
cm |
3 |
Infarcted lobe |
| 4421 |
Two |
175
g |
|
3 |
Old infarct |
| 4401 |
Two |
218
g |
27
cm |
3 |
Small infarct |
| 4058 |
Two |
120
g |
20
cm |
2 |
Infarct, stillb. |
| 4076 |
One |
130
g |
35
cm |
3 |
2 infarcts |
| 12657 |
Two |
Fetus
665 g |
20cm |
3 |
Villitis, died |
| 4145 |
Two |
35
g |
|
|
Abortion |
| 4195 |
One |
45
g |
|
|
Abortion |
| 3693 |
Two |
225
g |
18
cm |
3 |
Infarcts |
| 3754 |
Two |
159
g |
24
cm |
|
|
| 4168 |
Two |
145
g |
29
cm |
3 |
Infarct |
| 3117 |
Two |
227
g |
34
cm |
3 |
Living |
| 17095 |
Two |
125
g |
35
cm |
3 |
Infarcts |
5) Details of fetal/maternal
barrier
This is a mono-hemochorial relationship with the maternal blood surrounding
all villi, very similar to the placenta of other catarrhine monkeys. The
villi are covered mostly by syncytiotrophoblast; occasional cytotrophoblastic
cells lie beneath the syncytium and border the basement membrane. Syncytial
"knots" (or sprouts) occur frequently and are prominent at term.
They may detach and be swept away in the intervillous maternal blood, as
is normally the case in human placentas.
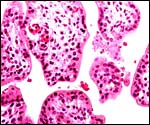 |
Immature villi of Hanuman placenta showing nucleated red blood cells in the fetal capillaries. |
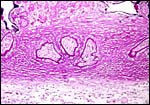 |
Base of implanted immature placenta, PAS stain. A broad villous connection (by extravillous trophoblast) anchors the villi to the fibrinoid of Nitabuch. |
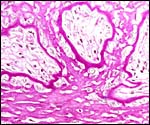 |
Same placenta at higher magnification. The avascular villous cores are covered with fibrinoid rather than trophoblast in the anchoring site. |
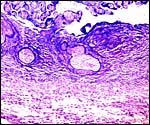 |
Same section in Giemsa stain. Note that there is virtually no decidual penetration by trophoblast. |
6) Umbilical cord
The umbilical cord of Hanuman langurs has normally three blood vessels,
2 arteries and one vein, and there is no patent allantoic duct. Occasionally,
single umbilical arteries occur, as is true of +/- 1% of human placentas.
Rarely, atrophied remnants of the allantoic duct may be found. There is
no surface squamous metaplasia. The variety of observed cord lengths in
my material will be seen in the preceding table. Other observations are
available in Bolk's contribution (1900) and the paper by Starck (1957).
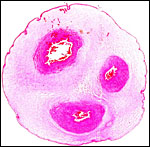 |
Cross section of typical umbilical cord of Hanuman langur. Vein is at top left. |
7) Uteroplacental
circulation
No special studies have been conducted in langurs, but it is reasonable
to assume this to be very similar to the vascular supply of rhesus monkeys
which is reviewed in that chapter. Occasional major maternal arteries in
the decidua basalis have infiltration with cytotrophoblast, as is shown
next. No really extensive modulation of maternal vessels occurs, however.
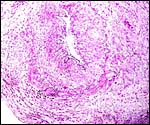 |
Large uterine decidual artery with minimal cytotrophoblast infiltration at its outside. |
8) Extraplacental
membranes
Burton (1980) stated that there is no decidual transformation in the young
placentations he observed in Presbytis obscura. There is, however,
extensive decidual transformation in mature placentas and I have been repeatedly
surprised at the large quantity of decidua capsularis in the membrane rolls
that were prepared from placentas. As is true in other cercopithecids, there
are no atrophying villi in the membrane rolls' decidua capsularis, as are
common features of ape and human placentas. The decidua capsularis is rich
in vessels and endometrial glands and it contains no evidence of trophoblast
infiltration. The amnion is thin and avascular. The epithelium is single-layered
and lacks squamous metaplasia. There is no allantoic sac. The yolk sac does
not persist.
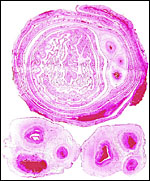 |
Membrane roll and sections of umbilical cords of Hanuman langur. The large vessels that cross between lobes are seen in the membrane roll. |
 |
A "membrane roll" of the free membranes between disks. Note the thick decidua capsularis and absence of atrophied villi. Amnion is outside. |
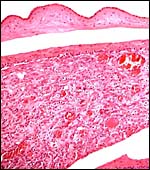 |
Membranes under higher magnification. Amnionic epithelium at top, with its avascular connective tissue, artifactually separated from the thin chorion which is underlain by a very small layer of cytotrophoblast. Most of the tissue is highly vascular decidua capsularis. |
9) Trophoblast
external to barrier
There is virtually no trophoblastic infiltration of the decidua basalis.
Only very few trophoblastic cells are found in the maternal vessels that
supply the intervillous space. Such an infiltration is commented upon only
in the early implantation stages of Presbytis obscura that Burton
(1980) was able to study.
10) Endometrium
As mentioned earlier, Burton (1980) found little decidual transformation
around the implantation site in early gestation. He remarked on the extensive
endometrial secretory activity adjacent, but not below the implantation
site and on the adjacent endometrial proliferation. In later gestation,
however, there is extensive decidual transformation that is especially
evident in the decidua capsularis of the membrane rolls.
11) Various features
There is no subplacenta.
12) Endocrinology
David & Ramaswami (1969) studied the menstrual cycle over many cycles
and found it to vary widely in length, with an average of 26.8 days. They
correlated this also with vulvar swelling and vaginal cornification. Progesterone
metabolites were ascertained in the urine of cycling and pregnant Hanuman
langurs by Shandilya et al. (1976a). Pregnanediol was the principal metabolite
during pregnancy and it rose a day after ovulation and reached its peak
on day 17 of the cycle. All three major estrogens were identified in the
urine of cycling and pregnant langurs (Shandilya et al., 1976b). All rose
significantly during gestation. Sialic acid levels were determined in
various reproductive organs of male and female langurs in an effort to
pinpoint the day of ovulation (Shandilya et at., 1977). It was not truly
effective in doing so. Ramaswami (1975), in a comprehensive review of
Hanuman langur reproductive biology, stated that there is a "pregnancy
Presbytis gonadotropin", but gave no further details and
levels are not yet known. Maston & Ruvola (2002) studied the genes
for the subunits of chorionic gonadotropin and found that man has 7 unique
clones, while Presbytis has 6. They also provided a tree for
the evolution of the LH ß duplication, the CGß gene, and derived
macacs and colobines as a group from a common ancestor.
13) Genetics
Presbytis spp. all have 44 chromosomes that are very similar among the various species; an example of Presbytis senex, the purple-faced langur, is shown following the one on Hanuman langurs (Hsu & Benirschke, 1977; see also Zneimer et al., 1979; Sharma & Kakati, 1967). A number of hybrids have been alluded to, but few have lived long enough to ascertain their possible fertility (Gray, 1972), at least this is not remarked upon. Sharma & Gupta (1973) suggested an autosomal/X-chromosomal translocation to have occurred in a specimen they studied; the photograph, however, is not convincing. Banding studies of two Presbytis species and macacs were published by Ponsa et al. (1983).
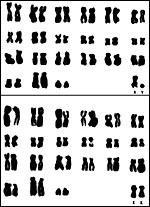 |
Karyotypes of male and female Presbytis entellus, 2n-44. |
 |
Karyotypes of male and female Presbytis senex. |
14) Immunology
I know of no studies other than the serologic surveys of experimental nature
reviewed in the next section.
15) Pathological features
There has been an active breeding colony of Hanumans at San Diego Zoo's
Wild Animal Park in the past. A significant problem there had been the occurrence
of intestinal perforation with peritonitis. It was attributed to the spines
of Acacia salignum, a plant the animals loved to consume; but they
suffered the peritonitis. Since discontinuing this diet, the problem disappeared
(Ensley et al., 1982). A few cases of stillbirth and abortion have occurred,
almost all of them associated with major placental infarction. Additionally,
villitis of unknown etiology ("VUE") has occurred, and we have
also seen acute villitis due to bacilli. VUE is a relatively common entity
that leads to growth restriction in human gestations and is currently not
fully understood (Benirschke & Kaufmann, 2000). Griner (1983) described
an insignificant cestode infestation, lobar pneumonia, and glomerulosclerosis
in his review of primate mortality at San Diego Zoo. Walker et al. (1973)
were able to show in Presbytis cristatus the induction of scrub
typhus with rickettsemia but no development of antibodies. They conjectured
that this tree-dwelling species was more susceptible than other monkeys
because of their lack of exposure to the vector. Mycobacterium bovis
was isolated from a dusky langur (Presbytis obscurus) with splenic
and intestinal lesions (Himes et al., 1982). The infection was acquired
from a group of newly imported fennec foxes that were housed in the same
facility. Benveniste & Todaro (1977) isolated a d-type retrovirus from
the genome of two langur species. David and Ramaswami (1967) reported right
renal hypoplasia in a female animal. Scott (1992) depicted acute myocarditis
(Fig. 7.2) and old pleurisy (Fig. 7.3) in langurs.
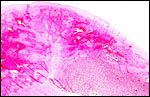 |
Typical, sharply circumscribed infarct (left) of term Hanuman langur placenta. |
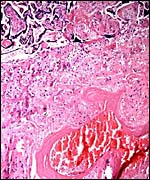 |
Decidua basalis with large maternal blood vessel showing fibrinoid replacement of its muscular wall. This is similar to minor changes of the decidual arteriopathy in cases of human pre-eclampsia. |
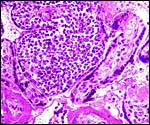 |
Chronic villitis ("VUE") of langur placenta. |
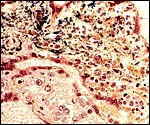 |
Acute necrotizing villitis in langur villi, Gram stain. Not black organisms and necrosis of villous core. Fetal demise. |
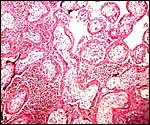 |
Acute, infectious villitis and "intervillositis" in abortus. |
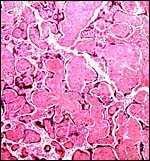 |
Complete hyalinization of villi in stillborn, aborted Hanuman langur gestation. |
 |
Complete cross section through the immature placenta of abortion specimen; no cause was identified. Chorion is above. |
16) Physiologic
data
Most important for the maintenance of langurs in captivity has been their
unusual (leaf-demanding) diet. This has been one reason for the difficulty
of maintaining many species of these langurs in captivity, and to keep them
healthy. Bauchop & Martucci (1968) made the most important observations
regarding this matter. While it had been known that the stomach contents
of the leaf-eaters may weigh as much as 15-20% of the body weight and that
their abdomens are often markedly protuberant, the true ruminant-like digestion
of leaves had not been appreciated before their study. They showed that
the stomach is partitioned and that there is a typical groove ("Magenstrasse"),
not unlike that of ruminants. But these monkeys do not ruminate. Huge numbers
of specific anaerobic, and few aerobic bacteria digest the leafy material
and produce volatile fatty acids, probably their most important energy requirement.
These authors also suggested from their experiments that fresh alfalfa was
eagerly consumed and that animals subsequently thrived on it after initial
starvation.
The maintenance under laboratory conditions has also been explored for the
silvered leaf monkey, Presbytis cristatus, by Walker et al. (1974).
Diarrhea, often due to salmonellosis, was their main concern, but methemoglobinemia
also occurred. Various hematologic values are available in that publication.
David & Ramaswami (1971) described in considerable detail the reproductive
organs of males and females and noted the similarity to the human anatomy
in several systems. The ovulation apparently is associated with considerable
hemorrhage and the blood is contained in the cystic corpus luteum (Ramaswami,
1975). He also stated that, unlike the rhesus monkey, the cervix is straight
and not spiraled.
17)
Other resources
Several cell strains of Hanuman langurs are available from CRES
at San Diego Zoo by contacting Dr. Oliver Ryder at oryder@ucsd.edu.
18) Other remarks - What additional
information is needed?
The reason for the frequent occurrence of placental infarcts is currently
not understood although minor vascular changes occur. Early stages of
implantation and placental development of Hanuman langurs are lacking.
Better knowledge of the endocrine status of gestation, especially of gonadotropins,
would be helpful.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society
of San Diego.
References
Bauchop, T. and Martucci, R.W.: Ruminant-like digestion of the langur
monkey. Science 161:698-700, 1968.
Benirschke, K.: Anatomical and functional differences in the placenta of primates. Biol. Reprod. 26:29-53, 1982.
Benirschke, K. and Kaufmann, P.: The Pathology of the Human Placenta, fourth edition. Springer-Verlag, NY, 2000.
Benveniste, R.E. and Todaro, G.J.: Evolution of primate oncornaviruses: An endogenous virus from langurs (Presbytis spp.) with related virogene sequences in other Old World monkeys. Proc. Natl. Acad. Sci. USA 74:4557-4561, 1977.
Bolk, L.: Beiträge zur Affenanatomie. I. Untersuchungen am schwangeren Uterus von Semnopithecus. Morphol. Jb. 28:565-623, 1900.
Burton, G.J.: Early placentation in the Dusky Leaf Monkey (Presbytis obscura). Placenta 1:187-195, 1980.
David, G.F.X. and Ramaswami, L.S.: Unilateral hypoplasia of the kidney of a female langur (Presbytis entellus entellus dufresne). Folia Primatol. 5:312-315, 1967.
David, G.F.X. and Ramaswami, L.S.: Studies on the menstrual cycles and other related phenomena in the langur (Presbytis entellus entellus). Fol. Primatol. 11:300-316, 1969.
David, G.F.X. and Ramaswami, L.S.: Reproductive systems of the North Indian langur (Presbytis entellus entellus Dufresne). J. Morphol. 135:99-107, 1971.
Ensley, P.K., Rost, T.L., Anderson, M.,
Benirschke, K., Brockman, D. and Ullrey, D.:
Intestinal obstruction and perforation caused by undigested Acacia
sp. leaves in langur monkeys. J. Amer. Vet. Med. Assoc. 181:1351?1354, 1982.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Gray, A.P.: Mammalian Hybrids. A Check-list
with Bibliography. 2nd edition.
Commonwealth Agricultural Bureaux Farnham Royal, Slough, England, 1972.
Griner, L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego, California, 1983.
Himes, E.M., Wendt, W.A., Luchsinger, D.W. and Jarnagin, J.L.: Mycobacterium bovis isolated from a dusky langur with granulomas in the intestine. JAVMA 181:1355-1357, 1982.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 10, Folio 517, 1977. Springer-Verlag, NY. (and Folio 349, 1973).
Maston, G.A. and Ruvola, M.: Chorionic gonadotropin has a recent origin within primates and an evolutionary history of selection. Mol. Biol. Evol. 19:320-335, 2002.
Nowak, R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press, Baltimore, 1999.
Ponsa, M., de Boer, L.E.M. and Egozcue, J.: Banding patterns of the chromosomes of Presbytis pyrrhus and P. obscurus. Amer. J. Primatol. 4:165-169, 1983.
Ramaswami, L.S.: Some aspects of the reproductive biology of the langur monkey Presbytis entellus entellus Dufresne. Sunder Lal Hora Medal lecture, Proc. Indian Natl. Sci. Acad. Part B1-30, 1975.
Roonwal, M.L. and Mohnot, S.M.: Primates of South Asia. Harvard University Pres, Cambridge, 1977.
Scott, G.B.D.: Comparative Primate Pathology. Oxford University Press, 1992.
Shandilya, L.N., Ramaswami, L.S. and Shandilya, N.: Progesterone metabolites in urine during menstrual cycle, pregnancy & puerperium in the Indian Hanuman langur Presbytis entellus entellus Dufresne. Indian J. Exp. Biol. 14:73-76, 1976a.
Shandilya, L.N., Ramaswami, L.S. and Shandilya, N.: Oestrogen metabolites in urine during the menstrual cycle, pregnancy and puerperium in the Indian hanuman langur (Presbytis entellus entellus). J. Reprod. Fertil. 47:7-11, 1976b.
Shandilya, L.N., Ramaswami, L.S. and Shandilya, N.: Sialic acid concentration in the reproductive organs, pituitary gland and urine of the Indian langur monkey (Presbytis entellus entellus). J. Endocrinol. 73:207-213, 1977.
Sharma, T. and Kakati, S.: Karyotype of Presbytis entellus entellus (Primate). Cytologia 32:111-116, 1967.
Sharma, G.P. and Gupta, C.M.: X-autosome translocation in the Indian langur - Presbytis entellus. Curr. Sci. 42:576, 1973.
Starck, D.: Ueber die Länge der Nabelschnur bei Säugetieren. Z. Säugetierk. 22:77-86, 1957.
Walker, J.S., Cadigan, F.C., Vosdingh, R.A. and Chye, C.T.: The silvered leaf-monkey of Malaysia, Presbytis cristatus: Disease model for human scrub typhus. J. Infect. Dis. 128:223-226, 1973.
Walker, J.S., Cadigan, F.C., Sirimanne, A.R. and Bass, J.B.: Silvered leaf-monkeys (Presbytis cristatus). I. Colony management, caging, diet, and biologic values. Lab. Anim. Sci. 24:290-298, 1974.
Zneimer, S., Kumamoto, A.T. and Benirschke, K.: Banding patterns of the chromosomes of two langur species. Pygathrix nemaeus and Presbytis entellus: A comparative study. Chromosome Inform. Serv. 26:19-22, 1979.