 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last
updated: March 10, 2004. |
Gazella granti (roosevelti)
Order - Artiodactyla
Family - Bovidae
1) General zoological data
The subfamily antilopinae has been confused ever since early attempts at classification (see Effron et al., 1976). This is further emphasized in the phylogenetic study of cytochrome b of some Bovidae by Matthee & Robinson (1999). Regrettably it did not include gazelle species. In a subsequent contribution (Matthee & Davis, 2001) their study showed that gazelles were always polyphyletic and suggested then that original diversification of Bovidae occurred in Eurasia. One of the largest of the "true gazelles" is the Grant's gazelle of East Africa (Kenya, Tanzania). Nine subspecies are listed by Wilson & Reeder (1992), of which this specimen is one. Nowak (1999) stated that the IUCN classified Grant's gazelle as a "conservation-dependent" species. The longevity of Gant's gazelles is 11 years, 11 months according to Jones (1993).
 |
Grant's gazelle at San Diego Wild Animal Park. |
Grant's gazelles have between 1 and 3 offspring and each weighs around 5 kg (Hayssen et al. 1993). The gestational length is given as ~ 6 months.
3) Implantation
Early stages of implantation have not been described but they are likely to be similar to other bovid species (Mossman, 1987).
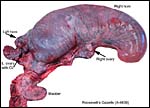 |
Pregnant uterus in right horn. |
This is a polycotyledonary, epitheliochorial placenta. The specimens shown here were in mid-pregnancy; the placentas had implanted in the right horn, while the corpus luteum was in the left ovary. Transuterine migration is not uncommon and the right horn preference of development is characteristic of many gazelles and some other species (Lee et al., 1977). While the first and better preserved specimen had exclusively right horn occupancy, the second fetus' placenta extended nearly symmetrically over both horns. There were 60 cotyledons arranged in 4 rows in one animal and 65 in the other. They were of the typically "convex" type (Mossman, 1987). They measured between 3 and 1 cm in diameters. The male fetuses were attached by an 8 and 10 cm cord, weighed 230 g and 241 g, and they measured 10 and 19 cm crown-rump length respectively. The second fetus was in early autolysis, obviously having died before the dam did. Both dams had disseminated infection secondary to pharyngeal abscesses.
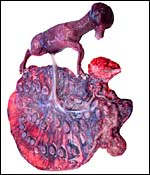 |
Male fetus attached to polycotyledonary placenta. |
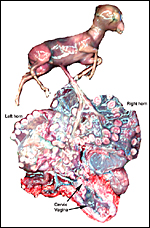 |
Macerated fetus in second gestation described with implantation in both horns. |
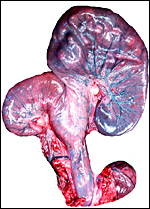 |
Unopened uterus of second gestation. While the fetus lay in the right horn, the placenta extended over both horns. Bladder below right. |
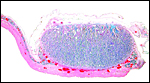 |
Typical cotyledon of this implanted placenta. Uterus below. |
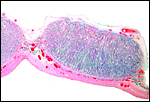 |
Two adjacent cotyledons with minimal endometrial glands between them. |
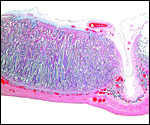 |
Another intercotyledonary region at right. |
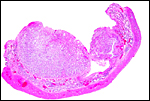 |
Another intercotyledonary region. |
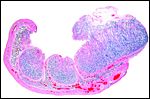 |
Groups of closely packed cotyledons. |
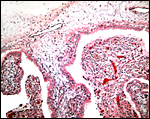 |
Chorionic surface (top) with caruncular interface (right). |
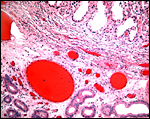 |
Base of cotyledon with the endometrium below. No trophoblastic invasion. |
5) Details of fetal/maternal barrier
The trophoblastic epithelium is cuboidal and interrupted by typical binucleate cells (Wooding, 1982; Atkinson et al., 1993) whose function presumably here also is the production of glycoproteins. ). These were studied in greater detail by Wooding et al. (1997) and are now considered to produce placental lactogen and other glycoproteins. In contrast to so many other ungulate placentas, these placentas had no areas of pigmentation or regions that could be interpreted as hemophagous organ.
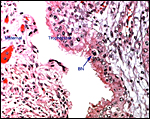 |
Maternal/fetal interface. The maternal epithelium (left) has undergone much degeneration, a post mortem artifact. BN=binucleate cell. |
6) Umbilical cord
These fetuses had cords measuring 8 and 10 cm in length that were covered with fine plaques of squamous metaplasia. They contained 4 large vessels, numerous delicate small vessels and had an allantoic duct. They had the characteristic site at mid-uterus and a mesometrial location (Mossman, 1987).
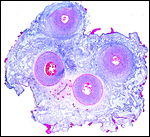 |
Umbilical cord of Grant gazelle with allantoic duct between vessels below. Note numerous foci of red surface squamous metaplasia. |
7) Uteroplacental circulation
This has not been studied.
8) Extraplacental membranes
There is a large allantoic duct filled with urine and lined by cuboidal epithelium. Its connective tissue membrane contains numerous small blood vessels.
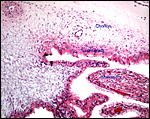 |
This is from the edge of the cotyledon and beginning of membranes. |
9) Trophoblast external to barrier
There are none.
10) Endometrium
The nonpregnant uterus has typical caruncles, arranged in four rows and here seen in the left nonpregnant horn. The endocervix was filled with tenacious mucus in both uteri.
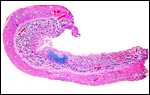 |
Section of the left, nonpregnant horn. Virtually no caruncles are seen. |
11) Various features
No other studies are known to me that are possibly relevant.
12) Endocrinology
The corpus luteum of this pregnancy was on the left, the right horn carried the fetus; thus, transmigration must have occurred. This is a common feature of many gazelle gestations.
13) Genetics
Grant's gazelles have 31 chromosomes in males, and 30 chromosomes in females. As in many other gazelles, a translocation between an autosome and X chromosome is present (Hsu & Benirschke, 1975; Effron et al., 1976; Gallagher & Womack, 1992) that explains the odd number of chromosomes in male gazelles. Hybrids have not been described. Arctander et al. (1996) studied the mtDNA control region of Grant's gazelles from different regions and found it to be highly divergent in different regions.
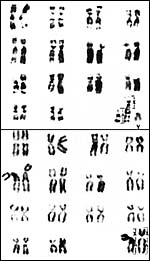 |
Karyotypes of male (top) and female (bottom) Grant's gazelles (from Hsu & Benirschke, 1975). |
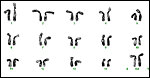 |
A more recent Karyotype of female Roosevelt 's gazelle from the San Diego Zoo. |
14) Immunology
I am not aware of any studies.
15) Pathological features
Griner (1983) reported on findings in 72 animals. By that time there had already been 77 births at San Diego , of which 24 had died neonatally and 6 were perinatal deaths. Trauma and a variety of infectious causes of death are listed in that review. Rumenitis with ulcers also occurred and one neonate had a cleft palate. Interestingly, four animals were icteric at birth for which the cause was not identified but possibly linked to a new male father. Baumeister et al. (1983) in a study of rotavirus infection of zoo animals found it primarily in young animals. Appleby et al. (1992) found the trichostrongylid nematode Cooperioides antidorca in intestinal nodules of animals in Kenya .
The remarkable feature of the second placenta described here is that, despite the fetus' significant maceration, the placental attachment had not suffered and there was very little degeneration of the villous tissue. Apparently, the endometrial perfusion of the caruncle was sufficient to maintain adequate structure, despite the absence of fetal circulation. The cord and its endothelia and allantoic epithelium had autolyzed.
16) Physiologic data
Pospisil et al. (1984) compiled various hematological parameters of Grant's gazelles and other species.
17) Other resources
Cell strains are available from CRES at the San Diego Zoological Society. We maintain many cell strains of Grant's gazelles at present. They can be obtained by contacting Dr. O. Ryder at oryder@ucsd.edu.
18) What additional information is needed?
Implantational stages and term placentas and their dimensions are unreported so far.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society of San Diego. I appreciate also very much the help of the pathologists at the San Diego Zoo.
References
Appleby, E.C., Gibbons, L.M. and Khalil, L.F.: Cooperioides antidorca from intestinal nodules in Gazella thomsoni and G. granti . J. Comp. Pathol. 107:295-303, 1992.
Arctander, P., Kat, P.W., Aman, R.A. and Siegismund, H.R.: Extreme genetic differences among populations of Gazella granti , Grant's gazelle in Kenya . Heredity 76:465-475, 1996.
Atkinson, Y.H., Gogolin-Ewens, K.J., Hounsell, E.F., Davies, M.J., Brandon , M.R. and Seamark, R.F.: Characterization of placentation-specific binucleate cell glycoproteins possessing a novel carbohydrate. J. Biol. Chem. 268:26679-26685, 1993.
Baumeister, B.M., Castro, A.E., McGuire-Rodgers, S.J. and Ramsay, E.C.: Detection and control of rotavirus infections in zoo animals. J. Amer. Vet. Med. Assoc. 183:1252-1254, 1983.
Effron, M., Bogart, M.H., Kumamoto , A.T. and Benirschke, K.: Chromosome studies in the mammalian subfamily Antilopinae . Genetica 46: 419-444, 1976.
Gallagher, D.S. and Womack, J.E.: Chromosome conservation in the Bovidae. J. Hered. 83:287-298, 1992.
Griner, L.A. : Pathology of Zoo Animals. Zoological Society of San Diego , San Diego , California , 1983.
Hayssen, V., van Tienhoven, A. and van Tienhoven, A.: Asdell's Patterns of Mammalian Reproduction: a Compendium of Species-specific Data. Comstock/Cornell University Press, Ithaca , 1993.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 9, Folio 439, 1975. Springer-Verlag , New York .
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Lee, S.Y., Mossman, H.W., Mossman, A.S. and del Pinto, G.: Evidence of specific nidation site in ruminants. Amer. J. Anat. 150:631-640, 1977.
Matthee , C.A. and Robinson, T.J.: Cytochrome b phylogeny of the family bovidae: resolution within the alcelaphini, antilopini, neotragini, and tragelaphini. Mol. Phylogenet. Evol. 12:31-46, 1999.
Matthee , C.A. and Davis , S.K.: Molecular insights into the evolution of the family Bovidae: A nuclear DNA perspective. Mol. Biol. Evol. 18:1220-1230, 2001.
Mossman, H.W.: Vertebrate Fetal Membranes. MacMillan, Houndmills, 1987.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Pospisil, J., Kase, F., Vahala, J. and Mouchova, I. : Basic haematological values in antelopes III. The Reduncinae and the Antelopinae. Comp. Biochem. Physiol. A 78:809-813, 1984.
Wilson , D.E. and Reeder, D.A.M.: Mammal Species of the World. A Taxonomic and Geographic Reference. 2 nd ed. Smithsonian Institution Press, Washington , DC , 1992.
Wooding, F.B.: The role of the binucleate cell in ruminant placental structure. J. Reprod.
Fertil. Suppl. 31:31-39, 1982.
Wooding, F.B., Morgan, G. and Adam, C.L.: Structure and function in the ruminant synepitheliochorial placenta: central role of the trophoblast binucleate cell in deer. Microsc. Res. Tech. 38:88-99, 1997.