 |
| (Clicking
on the thumbnail images will launch a new window and a larger version
of the thumbnail.) |
| Last updated: July 3, 2004. |
Taurotragus oryx (pattersonianus)
Order Artiodactyla
Family Bovidae
1) General Zoological Data
There are two species of eland, the common eland ( Taurotragus oryx pattersonianus ) described here, and the Giant or Derby eland, T. derbianus. Both are large animals from Africa and range from Ethiopia south (Nowak, 1999). Tragelaphus (from Gr. Tragos = he-goat, and elaphos = deer). They are widely represented in numerous zoos and do well in captivity. A large captive herd was established in Askania Nova ( Russia ) and was used for milk production. In contrast to greater kudus, the females possess horns, though less spiraled than in male kudus. The evolutionary relationships of various bovid taxa were described with cytochrome b mtDNA data by Hassanin & Douzery (1999) and Matthee & Robinson (1999).
 |
Male Patterson's eland at San Diego Zoo's Wild Animal Park. |
 |
Female Patterson's eland with young at San Diego Zoo's Wild Animal Park. |
2) General Gestational Data
The estrous cycle of 21-26 days is followed by a 1-3 days period of estrus; gestation lasts 254-277 days (Nowak, 1999; see also Mentis, 1972: average 276 days). A single calf is normally born weighing 22-36 kg. Longevity is maximally 23 ½ years, usually <18 years (Mentis, 1972). Occasional twins have been born in San Diego (1901). Early stages of implantation are not recorded. The maternal weight at calving is 550-650 lbs. (Mentis, 1972). The placental weight of my specimen was 1,100 g.
3) Implantation
Early stages of implantation are not recorded. Implantation occurs on flat caruncles, with a typical epitheliochorial, cotyledonary type of placenta evolving. Hradecky (1983) described implantation in right and left uterine horns. He described four rows of caruncles and observed cotyledons to develop in the lesser horn as well.
4) General Characterization of the Placenta
This is a typical polycotyledonary placenta whose cotyledons were arranged in four rows. The term, delivered organ I had available for study weighed 1,100 g and had only 25 cotyledons; it may have been incompletely submitted. Hradecky (1983) found 142 and 151 cotyledons in two placentas available to him, and Turner (1879) had more than 100 cotyledons. In addition, an attached placenta was studied.
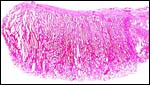 |
Margin of one eland cotyledon from a delivered placenta. |
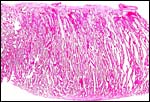 |
Another section of the villous tissue of a delivered eland placenta. |
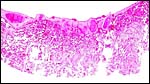 |
Another section of villous tissue from a delivered placenta showing a better fetal surface with fetal blood vessels. There is no maternal tissue. |
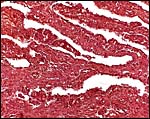 |
Eland villous tissue interlaced with maternal septa. |
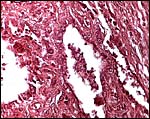 |
Trophoblastic surface of villi. |
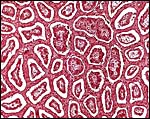 |
The network represents the maternal tissue of an implanted eland placenta; villi in between. |
5) Details of fetal/maternal barrier
This is an epitheliochorial relation between trophoblast and the intricate network of maternal septa. This is well shown in the cross section of cotyledons and the structures have been detailed in a study of 18 eland placentas by Hradecky et al. (1987). Finely divided yellow granular material is present in some enlarged subchorionic trophoblastic cells. It is uncertain whether this represents the hemophagous tissue, as these cells are absent in my placenta from an admittedly immature greater kudu specimen. Typical binucleate cells are present but not abundant. Microvilli extend from the trophoblastic surface. The uterine epithelium in this specimen is single-layered and intact, albeit somewhat autolyzed.
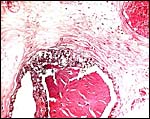 |
Subchorionic trophoblast of a term eland placenta contains small yellow-brown pigment granules. |
6) Umbilical cord
The umbilical cord has four blood vessels and a large allantoic duct. Foci of squamous metaplasia cover the surface. Many small blood vessels are present in addition to the four large ones.
7) Uteroplacental circulation
No details have been described.
8) Extraplacental membranes
There is a large allantoic sac lined by flat epithelium and abundantly vascularized. The amnion was smooth and without areas of squamous metaplasia. Hippomanes were absent. The trophoblast extends between the cotyledons on the outside of the membranes.
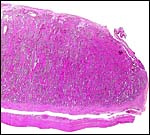 |
Margin of implanted cotyledon with intercotyledonary membrane shown at top right. The myometrium is artifactually detached, below. |
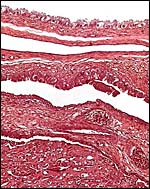 |
Edge of cotyledon with trophoblast of intercotyledonary membrane above. |
9) Trophoblast external to barrier
There is no infiltration of the basal endometrium.
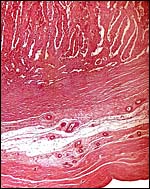 |
Implanted eland placenta near term. Myometrium below. |
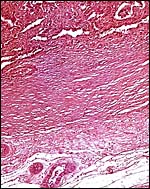 |
The maternal septa of the endometrium are seen to extend from the uterine wall below. |
10) Endometrium
No true decidua was found. Details of caruncles and their relationship to implanted cotyledons can be found described in considerable detail by Hradecky et al. (1988).
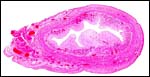 |
Fetal uterus of eland with pale caruncles extending into the endometrial cavity. |
11) Various features
There is neither a subplacenta nor are there metrial glands. The uterine morphology and that of the cervix have been described by Hradecky (1982) for this antelope and for other related bovid species. The same author (1984) also published his findings of the composition of the allantoic and amniotic fluids and found them to be highly variable.
12) Endocrinology
The estrous cycle is 21-26 days (Nowak, 1999) and estrus lasts 1-3 days. The role of hormone production (placental lactogen) by the trophoblastic binucleate cells was explored by Wooding (1982). Atkinson et al. (1993) explored the glycoprotein production in ovine placentas by these cells. The fetal ovary shows no evidence of hormonal stimulation. The adrenal glands of adults and newborns are not unusual in their structures when compared with other mammals. Aspect of endocrine control of mammary growth and function are considered by Forsyth (1986).
13) Genetics
Chromosome studies were studied since the 1970s and the findings referred to in the references of Hsu & Benirschke (1971). Males have 31 chromosomes with a fused Y-chromosome (t Y/14), females have 32 chromosomes as shown below. Studies with tritiated thymidine have indicated that the large acrocentric element is the X-chromosomes, fused with a small autosome in the males. Hence their chromosome number is one less than that of females. (See also Wurster, 1972).
 |
Karyotypes of male and female elands. |
Although mating with cows apparently occurs readily, hybrids have not been produced with cattle. Hybrids with greater kudu, however, are possible and the male one hybrid studied in detail at least was sterile (Gray, 1972; Jorge et al., 1976). This animal and its parents are shown below. Since both species have the same chromosome number, the sterility may be due to the different sizes and shapes of chromosomes from putative inversions, as detailed in the latter contribution. Gallagher & Womack (1992) also studied the translocations of many bovid species, including the greater kudu and suggested that speciation may have been the result of translocation with subsequent isolation of resultant karyotypically rearranged individuals (see also Wurster et al., 1972).
In addition, Petit et al. (1994/5) studied the relationships of these African animals. Both views are presented at the end in a pedigree of possible developments over time. The finding of translocations in both sex chromosomes contrasts with that of many other African ungulates that have only an X/A translocation, but possess a normal Y chromosome. These species thus have different numbers in males and females (Benirschke et al., 1980). Other views of evolutionary relationships, resulting from mtDNA studies, were presented by Matthee & Robinson (1999) and Hassanin & Douzery (1999).
 |
Eland (m) x kudu (f) hybrid (Jorge et al., 1976). |
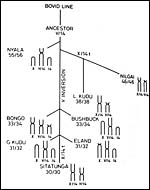 |
Possible relationship of tragelaphines (from Benirschke et al., 1980). |
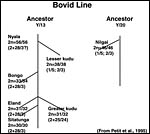 |
This putative phylogeny is arranged by Petit (1995) according to cytogenetic information with chromosome numbers (2n=males/females) and the types of fusions as arranged according to bovid karyotype agreement. |
14) Immunology
I am not aware of any publications.
15) Pathological features
Griner (1983) described his experience with autopsies of 20 eland only minimally. Trauma was the principal cause of mortality; one eland died with bronchial adenocarcinoma; infectious diseases are not listed. The hybrid shown below was euthanized because of its sterility.
16) Physiologic data
Pospisil et al. (1984) published hematological values of hippotraginae and tragelaphinae, including those of the greater kudu. Hagey et al. (1997) described the changes in bile acid composition with age a many bovids.
17) Other resources
Cell strains of this and several other animals as well as related tragelaphines are available from CRES by contacting Dr. Oliver Ryder at: oryder@ucsd.edu .
18) Other remarks What additional Information is needed?
Early stages of implantation are not on record and more endocrine data are needed.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society of San Diego.
References
Atkinson, Y.H., Gogolin-Ewens, K.J., Hounsell, E.F., Davies, M.J., Brandon , M.R. and Seamark, R.F.: Characterization of placentation-specific binucleate cell glycoproteins possessing a novel carbohydrate. J. Biol. Chem. 268:26689-26685, 1993.
Benirschke, K., Ruedi, D., Muller, H., Kumamoto , A.T. Wagner, K.L. and Downes, H.S.: The unusual karyotype of the lesser kudu, Tragelaphus imberbis. Cytogenet. Cell Genet. 26:85-92, 1980.
Forsyth, I.A.: Variation among species in the endocrine control of mammary growth and function: the roles of prolactin, growth hormone, and placental lactogen. J. Dairy Sci. 69:886-903, 1986.
Gallagher, D.S. and Womack, J.E.: Chromosome conservation in the Bovidae. J. Hered. 83:297-298, 1992.
Gray, A.P.: Mammalian Hybrids. A Check-list with Bibliography. 2 nd edition.
Commonwealth Agricultural Bureaux Farnham Royal, Slough , England , 1972.
Griner, L.A. : Pathology of Zoo Animals. Zoological Society of San Diego , San Diego , California , 1983.
Hassanin, A. and Douzery, E.J.: The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome b gene. Mol. Phylogenet. Evol. 13:227-243, 1999.
Hradecky, P.: Uterine morphology in some African antelopes. J. Zoo An. Med. 13:132-136, 1982.
Hradecky, P.: Placental morphology in African antelopes and giraffes. Theriogenology 20:725-734, 1983.
Hradecky, P.: Composition of the fetal fluids in African antelopes. Theriogenology 22:657-665, 1984.
Hradecky, P., Benirschke, K. and Stott, G.G.: Implications of the placental structure compatibility for interspecific embryo transfer. Theriogenology 28:737-746, 1987.
Hradecky, P., Mossman, H.W. and Stott, G.G.: Comparative development of ruminant placentomes. Theriogenology 29:693-729, 1988.
Hsu, T.C. and Benirschke, K.: An Atlas of Mammalian Chromosomes. Vol. 6, Folio 295, 1971; Springer-Verlag, N.Y.
Jorge, W., Butler , S. and Benirschke, K.: Studies on a male eland x kudu hybrid. J. Reprod. Fert. 46 :13-16, 1976.
Matthee , C.A. and Robinson, T.J.: Cytochrome b phylogeny of the family Bovidae: Resolution within the Alcelaphini, Antilopini, Neotragini, and Tragelaphini. Mol. Phylogenet. Evol. 12:31-46, 1999.
Mentis, M.T.: A review of some life history features of the large herbivores of Africa . The Lammergeyer 16:1-89, 1972.
Nowak, R.M.: Walker 's Mammals of the World. 6 th ed. The Johns Hopkins Press, Baltimore, 1999.
Petit, P., Vermeesch, J.R., Marynen, P. and de Meurichy, W.: Comparative cytogenetic study in the subfamily Tragelaphinae. Proc. 11 th Europ. Coll. Cytogenet. Domest. Anim. Pp. 109-113, 1994/5.
Pospisil, J., Kase, F., Vahala, J. and Mouchova, I. : Basic haematological values in antelopes II. The Hippotraginae and the Tragelaphinae. Comp. Biochem. Physiol. A 78:799-807, 1984.
Turner, W.M.: On the fetal membranes of the eland ( Oreas canna ). J. Anat. Physiol. 14:241-243, 1879.
Wooding, F.B.: The role of the binucleate cell in ruminant placental structure. J. Reprod. Fertil. Suppl. 31:31-39, 1982.
Wurster, D.H.: Sex-chromosome translocations and karyotypes in bovid tribes. Cytogenetics 11:197-207, 1972.