 |
|
(Clicking
on the thumbnail images below will launch a new window and a larger
version of the thumbnail.)
|
Madoqua kirkii
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
There are at least five "good" species of these small antelopes. Guenther's dik-dik and Kirk's dik-dik are the most commonly examined dik-diks from Africa in zoological gardens, and may be difficult to distinguish (Kingswood & Kumamoto, 1997). All dik-diks are eastern African species, except for Kirk's variety which is also represented in the southeast of Africa, in Namibia. They form a dissociated population from the eastern form. Although now grouped with "Madoqinae", the Kirk's dik-dik was once also identified as Rhynchotragus because of their elongated snouts; further taxonomic features have been listed by Kingswood & Kumamoto (1997). [Madoqua is said to be from the Amharic and means: small antelope. Gotch, 1979]. Kirk's dik-dik has perhaps as many as six subspecies, and a seventh in southwest Africa (Haltenorth & Diller, 1980). Some animals are also described from Arabia and are there endangered (Habibi, 1986). A very detailed review of these species was undertaken by Kingswood & Kumamoto (1997). Adults weigh between 4 and 5 kg.
Dik-diks have a long lineage, dating back some 12 MYA. They are part of Neotragini which are included in the detailed study of mtDNA cytochrome b study of Matthee & Robinson (1999). They found this group to be polyphyletic, and only Guenther and Kirk's dik-diks are more closely related.
Sexual maturity is attained at age 6-8 months (Mentis, 1972). Longevity in captivity is over 14 years (Jones, 1993), but much less in the wild. Several breeding colonies exist in zoological gardens.
 |
Kirk's dik-dik at San Diego Zoo. |
 |
Guenther's dik-dik at San Diego Zoo. |
The length of gestation is from 170-174 days (Kellas, 1955), and implantation is generally in the right uterine horn, mesometrially. The sex ratio of captive-born animals was: M169/F134 (Bent & Reason, 1998). The litter size is usually singletons weighing 560-680 g.
3)
Implantation
Kirk's dik-dik placentation occurs usually in the right uterine horn.
It develops a polycotyledonary, epitheliochorial placenta that is not
invasive. Very few implanted placentas have been described, however, and
more information on the site of attachment is needed. The best observations
were made by Wislocki (1941). He had received an alcohol-fixed pregnant
uterus from a hunter and described it in great detail. Unlike any other
ruminant, his observations of a cotyledonary arrangement on "a stalk"
are surely erroneous, however, despite the pictures that accompany the
paper. It is my assumption that the stalks shown by him were produced
by the alcohol fixation of the specimen. Alcohol tends to contract the
muscle (myometrium) much more than the softer tissues and thus a stalk
may have been produced artefactually. Besides, Strahl (1911) had earlier
seen a complete, pregnant uterus and did not find this most unusual arrangement.
I, therefore, believe that the stalks are a fixation artifact. Wislocki
admits that his specimen suffered from some post mortem change. In addition,
he counted "about 80 cotyledons", but depicts far less, more
like the numbers here presented, around 40-50. The fixation must also
be the reason why his cotyledons had such pronounced convex shape, when
I and others found them to be flat in dik-diks.
4)
General Characterization of the Placenta
The placenta of dik-diks is polycotyledonary and epithelio-chorial. I
have been able to examine only this one term placenta in some detail.
It was a delivered placenta and, thus, it did not have any maternal tissue
in between the villi. The placenta had been peeled away from the caruncles
at delivery in the usual manner. No maternal tissue accompanied it. It
is here depicted and had 41 cotyledons, weighed 47 g, and measured 44x11x0.1
cm. The cotyledons varied from 2x0.1 to 1x0.1 cm in dimensions. The placenta
mentioned in Wislocki's report was described to have "about 80 cotyledons"
(1941). The cord inserts in the center.
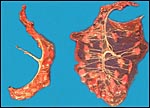 |
Term placenta of Kirk's dik-dik with central cord insertion and 41 cotyledons. |
The villi of this animal's placenta are not so very different from those of other antelopes. Wislocki felt that insufficient material had been collected as yet and from mostly inadequately fixed uteri so as to allow categorization of the structure of the villi. Besides, most authors tend to section their material somewhat differently, adding to the confusion. The trophoblast that covers the villi is cuboidal, single-layered and has occasionally binucleated trophoblastic cells, as is so typical for ruminants. The trophoblast has a fine brush border of microvilli. Some minor pigment granules are found here and there. My specimen did not have the maternal hematomas described by Strahl (1911), nor did the specimen studied by Wislocki (1941).
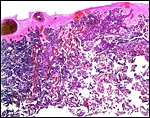 |
Section of delivered, term dik-dik placenta. Obviously, there are no maternal tissues in delivered placentas of dik-diks, as they remain in the uterus. |
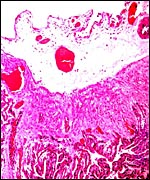 |
Surface of this delivered placenta to show course of connective tissue and blood vessels. |
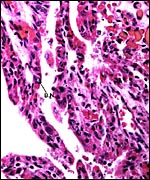 |
Higher magnification with villi having cuboidal trophoblast and occasional binucleate cells (B.N.). Fetal capillaries bulge into the trophoblast. |
The umbilical cord measured 8 cm in length and 0.3 cm in thickness. It contained four vessels, two arteries and two veins. In addition, there is the usual allantoic duct and numerous small blood vessels, most numerous adjacent to the duct. There were no spirals. Wislocki's observation of the immature gestation is that the umbilical cord was only 6 cm long, and it was studded with tiny epithelial protrusions (caruncles).
7)
Uteroplacental circulation
I know of no physiologic studies of the placentation, other than the microscopic
observations of blood supply to the endometrial caruncles and the remarks
by Wislocki (1941) that endometrial trabeculae and fetal villi had a rich,
similar vascularization. He also mentioned that fetal villous capillaries
often indent the trophoblast.
8)
Extraplacental membranes
In Wislocki's description (1941) of a complete uterus, the allantoic sac
nearly filled the left uterine horn, while the fetus was in the right
side, within its amnion (1941). No metaplastic nodules or squamous metaplasia
were present on this or in Wislocki's specimen. In the term placenta I
examined, there had been meconium staining. As a result, the amnion is
markedly degenerated, as is the case in human placentas following meconium
discharge. No pigmented macrophages were present within the membranes,
although meconium was present on the amnionic surface. The amnion is avascular;
numerous blood vessels are in the allantoic membrane and allantochorion.
The outer surface of membranes, the portion extending between the cotyledons,
is covered with tall, columnar trophoblast. Beneath this, mostly over
what must have been areolae in situ, is much cellular debris, as
is also mentioned by earlier observers. Maternal blood extravasation was
not present, a feature described by Strahl (1911) but not found by Wislocki
(1941). There were no hippomanes.
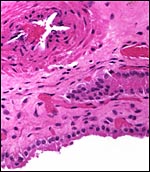 |
Chorioallantoic membrane between the cotyledons. It is covered (on the outside, below) with columnar trophoblast that has prominent microvillous surfaces. Allantoic blood vessel above. |
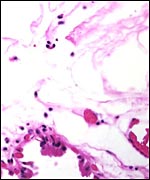 |
"Membranes" between cotyledons showing the allantoic sac below and the amnion above. It is degenerated because of the meconium discharge. |
There is no extravillous trophoblast. The villous surface, while apposed to the endometrium, does not invade the uterus at any place.
10)
Endometrium
The endometrium of ungulates undergoes considerable changes during gestation,
although it does not acquire the decidual transformation of primates,
not is the placenta detached in a decidual plane. Rather, the villi are
peeled out of their caruncular crypts. It is true, however, as remarked
by Wislocki (1941) that the endometrial glandular surface of the cisternae
into which the villi project, is covered by a peculiar epithelium. While
this could not be observed, of course, in this delivered placenta, it
is the case in other ruminant specimens I have had (see slender-horned
gazelle, for instance). The epithelium is flat, occasionally bunched,
and Wislocki considered it to be possibly syncytial. Moreover, because
of the apparent absence of some epithelium in the H&E preparations,
he stated that one should perhaps consider this placenta as being syndesmochorial,
rather than epithelio-chorial. Until electron microscopic study is undertaken,
I would not favor such an assignment. At the base of the cotyledons, the
endometrial stroma is dense, has large nuclei and occasional giant cells.
But, these are not trophoblastic and their nature and origin are unknown.
This is discussed extensively by Wislocki (1941). Despite the debris that
also exists in this region, I hesitate to refer to this as a "subplacenta',
as exists in carnivora.
11)
Various features
I do not deem the changes just described that occur at the base of cotyledons
sufficiently structured to speak of a "subplacenta".
12)
Endocrinology
I know of no endocrine studies in this species. Because of the binucleate
cells, one must assume that the placenta makes relaxin.
13) Genetics
The dik-diks have complex chromosomal arrangements. Animals with 2n=46
to 2n=48 seem to be the usual, but an animals with 2n=49 has existed as
well. Some possess 47 chromosomes with X/A translocation (Benirschke &
Kumamoto, 1987; Kumamoto et al., 1994; 1995). The two common "cytotypes"
(46 and 48 chromosomes) are sufficiently different that hybrids are sterile
(Kingswood & Kumamoto, 1997). Many zoos are now known to harbor hybrids
between different cytotypes, with anomalous chromosome numbers and causing
unexplained sterility. Moreover, examination of the testes of such hybrids
has shown subfertility or lack of spermatogenesis. In addition, hybrids
between Kirk's and Guenther's dik-dik exist and these are also infertile
(Ryder et al., 1989). Albinism has been described in the past (Roosevelt
& Heller (1915).
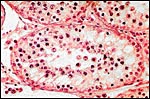 |
Testis of mature animal - hybrid between cytotypes A and B, showing absent spermatogenesis. |
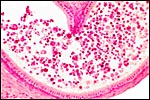 |
Epididymis of hybrid between cytotypes A and B showing absent spermatozoa. |
I know of no studies.
15)
Pathological features
Numerous ectoparasites occur, and pneumonia has resulted from cold exposure.
Cysticercosis infection was reported to occur in the Serengeti Park by
Sachs & Sachs, 1968. Few other diseases are listed by Kingswood &
Kumamoto (1997).
16)
Physiologic data
Numerous studies have been done in dik-diks that concern their water metabolism,
heat regulation and other aspects that explain their ability to withstand
the severe climatic conditions to which they are exposed. These are accessible
though the review by Maloy et al. (1988).
17)
Other resources
Cell lines of many dik-dik species are available from CRES
at the San Diego Zoo by contacting Dr. Oliver Ryder at: oryder@ucsd.edu.
18)
Other remarks - What additional Information is needed?
More data on placental weight and implanted placentas are needed. Importantly,
are the cotyledons on stalks or not?
Acknowledgement
I appreciate very much the help of the pathologists at the San Diego Zoo.
References
Benirschke, K. and Kumamoto, A.T.: Challenges of artiodactyl cytogenetics.
La Kromosoma II-45:1468-1478, 1987.
Bent, N. and Reason, R.: A preliminary study of sex ratios in captive-born ruminants. Int. Zoo Ybk. 36:223-228, 1998.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Habibi, K.: Arabian ungulates - their status and future protection. Oryx 20:100-103, 1986.
Haltenorth, T. and Diller, H.: A Field Guide to the Mammals of Africa including Madagascar. Collins Sons, Glasgow, 1980.
Jones, M.L.: Longevity of ungulates in captivity. Int. Ybk. 32:159-169, 1993.
Kellas, L.M.: Observations on the reproductive activities, measurements and growth of the dik-dik (Rhyncotragus kirkii thomasii Neumann). Proc. Zool. Soc. London 124:751-784, 1955.
Kingswood, S.C. and Kumamoto, A.T.: Madoqua kirkii. In, Mammalian Species # 569, pp. 1-10, Amer. Soc. Mammalogists, 1997.
Kumamoto, A.T., Kingswood, S.C. and Hugo, W.: Chromosomal divergence in allopatric populations of Kirk's dik-dik, Madoqua kirki (Artiodactyla, Bovidae). J. Mammal. 75:357-364, 1994.
Kumamoto, A.T. and Kingswood, S.C.: Description of a fourth cytotype for Kirk's dik-dik, Madoqua kirki (Artiodactyla, Bovidae). Cytogenet. Cell Genet. 71:400 (abstr.), 1995.
Maloy, G.M.O., Rugangazi, B.M. and Clemens, E.T.: Physiology of the dik-dik antelope. Mini Review. Comp. Biochem. Physiol. 91A:1-18, 1988.
Matthee, C.A. and Robinson, T.J.: Cytochrome b phylogeny of the family bovidae: Resolution within the alcelaphini, antilopini, neotragini, and tragelaphini. Mol. Phylog. Evol. 12:31-46, 1999.
Mentis, M.T.: A review of some life history features of the large herbivores of Africa. The Lammergeyer 1-89, 1972.
Roosevelt, T. and Heller, E.: Life-histories of African game animals. John Murray, London, 2:421-798, 1915.
Ryder,
O., Kumamoto, A.T., Durrant, B.S., and Benirschke, K.: Chromosomal divergence
and reproductive isolation in dik-diks. In, Speciation and its Consequences.
Chapter 9, pp. 208-225. D. Otte and J.A. Endler, eds. Sinauer Assoc. Sunderland,
Maryland, 1989.
Sachs, R. and Sachs, C.: A survey of parasitic infestation of wild herbivores
in the Serengeti region of Northern Tanzania and the Lake Rukwa region
in Southern Tanzania. Bull. Epizoot. Dis. Afr. 16:455-472, 1968.
Strahl, H.: Zur Kenntnis der Wiederkäuerplacentome. Anat. Anz. 40:259-2264, 1911.
Wislocki, G.B.: The placentation of an antelope (Rhynchotragus kirkii nyikae Heller). Anat. Rec. 81:221-241, 1941.