| |
13)
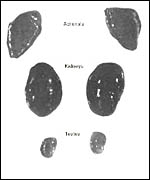
Genetics
The karyotype of these two species has been confusing and it is very different
from, and more complex than that of three-toed sloths. Especially the sex-determining
chromosomes have been a problem in their clear delineation. Because it is
difficult to determine the gender of two-toed sloths externally, Murata
& Masuda (1996) developed a technique that employs DNA from hair for
sex assignment. They amplified the SRY region and were able to determine
maleness in newborns.
Several authors have studied the chromosomes of Hoffmann's sloths and found
initially 2n=49, with the Y-chromosome translocated to a small autosome.
Females, however, also have only 49 chromosomes, with a single X-chromosome.
Two animals (perhaps representing subspecific hybrids) were supplied to
us for study by Meritt (1977). Both, one male and one female, had 50 chromosomes.
Jorge et al. (1985) found further variations, and identified animals with
chromosome numbers between 2n=49 and 51 for Hoffmann's sloths. The animals
that they studied were from specified regions in South America.
The situation is different in C. didactylus. Sonta et al. (1980)
found the two sexes to have 53 chromosome and a somewhat similar XO female
sex determination. They found a translocated Y/A as well. The most comprehensive
study undertaken on this species is that of Jorge et al. (1985). They found
six different karyotypes in animals from specified, different regions in
South America. Their chromosome numbers varied from 53 to 64 elements. It
is thus certain that there may well be subspecies/species differences in
different regions of South America that require much more intensive study.
No interspecific hybrids have been produced, but Meritt (1977) referred
to probable intersubspecific crosses in his study. Furthermore, the
very unusual chromosomal distribution found in these sloths suggests that
the nominated subspecies may actually be good species. Much further work
is needed before the situation is resolved.
14) Immunology
I know of no studies.
15)
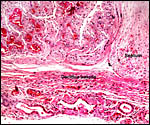
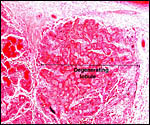
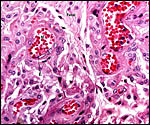
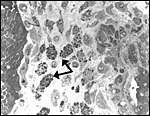
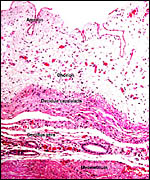
Pathological features
Seymour et al. (1983) accounted for the viruses isolated from sloths.
Griner (1983) found infection and malnutrition to be problems in captivity.
16) Physiologic data
Meritt (1973) provided dietary details for feeding sloths in captivity,
and Bush & Gilroy (1979) gave data on how to bleed sloths, and provided
hematologic data at the same time. Gilmore et al. (2000) reviewed the
general physiology of both types of sloths. In a later contribution (2001)
they updated this information and gave data on infectious organism harbored
by sloths. Clinical problems encountered in captivity were described by
Diniz & Oliveira (1999). Effective means of immobilization of Hoffmann's
sloths were described by Meritt (1972, 1974, 1985). The same observer
also made observations that allow identification of recently parturient
females in a group of animals (Meritt, 1976). Since parturition had not
been witnessed and mothers were difficult to identify, he used vaginal
orifice swabs and Hemastix to identify the mother of newborns.
17) Other resources
Cell strains are kept of some animals at CRES
in the San Diego Zoo and can be obtained by contacting Dr. Oliver Ryder
at: oryder@ucsd.edu
18)
Other remarks - What additional Information is needed?
Endocrine studies are badly needed. The nature of the xenarthran fetal
adrenal glands is of particular interest. Accurate data on the length
of gestation of both species are also essential, as they differ so much
in the few reports available. Are the chromosome differences indicative
of "good" species? Are "hybrids" among them infertile?
Acknowledgement
The animal photographs in this chapter come from the Zoological Society
of San Diego. I appreciate also very much the help of the pathologists
at the San Diego Zoo.
References
Becher, H.: Zur Kenntnis der Placenta vom Bradypus tridactylus.
Z. Anat. Entwicklungsgesch. 61:114-136, 1921.
Benirschke,
K. and Powell, H.C.: On the placentation of sloths. Pp. 237-241, In, Montgomery,
G.G., ed.: The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas.
Smithsonian Institution, Washington, 1985.
Bush,
M. and Gilroy, B.A.: A bleeding technique from nonpalpable vessels in
anesthetized two-toed sloths (Choloepus didactylus) - plus hematologic
data. J. Zoo Anim. Med. 10:26-27, 1979.
Delsuc,
F., Catzeflis, F.M., Stanhope, M.J. and Douzery, E.J.: The evolution of
armadillos, anteaters and sloths depicted by nuclear and mitochondrial
phylogenies: implications for the status of the enigmatic fossil Eurotamandua.
Proc. Roy. Soc. Lond. B Biol. 268:1605-1615, 2001.
Diniz,
L.S. and Oliveira, P.M.: Clinical problems of sloths (Bradypus sp.
and Choloepus sp.) in captivity. J. Zoo Wildl. Med. 30:76-80, 1999.
Eisenberg,
J.F. and Maliniak, E.: Reproduction of the two-toed sloth Choloepus
hoffmanni in captivity. Amer. Soc. Mamm. Abstr. Techn. Pap. 58th Ann.
Meet. 41-42, 1978 (cited by Nowak, 1999).
Eisenberg,
J.F. and Maliniak, E.: Maintenance and reproduction of the two-toed sloth
Choloepus didactylus in captivity. Pp. 327-331, in, The Evolution
and Ecology of Armadillos, Sloths, and Vermilinguas. G.G. Montgomery,
ed. Smithsonian Institution, Washington, 1985.
Gilmore,
D.P., da-Costa, C.P. and Duarte, D.P.: An update on the physiology of
two- and three-toed sloths. Brazil. J. Med. Biol. Res. 33:129-146, 2000.
Gilmore,
D.P., da Costa, C.P. and Duarte, D.P.: Sloth biology: an update on their
physiological ecology, behavior and role as vectors of arthropods and
arboviruses. Braz. J. Med. Biol. Res. 34:9-25, 2001.
Gotch,
A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset,
1979.
Greenwood,
A.D., Castresana, J., Feldmaier-Fuchs, G. and Paabö, S.: A molecular
phylogeny of two extinct sloths. Mol. Phylogenet. Evol. 18:94-103, 2001.
Griner,
L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego,
California, 1983.
Heuser,
C.H. and Wislocki, G.B.: Early development of the sloth (Bradypus griseus)
and its similarity to that of man. Contrib. Embryol. Carnegie Inst. Washington
25:1-13, 1935.
Jorge,
W., Meritt, D.A. and Benirschke, K.: Chromosome studies in edentates.
Cytobios 18:157-172, 1978.
Jorge,
W., Orsi-Souza, A.T. and Best, R.: The somatic chromosomes of Xenarthra.
Pp.121 in, The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas.
G.G. Montgomery, ed. Smithsonian Institution, Washington, 1985.
McCrane,
M.P.: Birth, behavior and development of a hand-reared two-toed sloth.
Int. Zoo Ybk. 6:153-163, 1966.
Meritt, D.A.: Edentate immobilisation at Lincoln Park Zoo, Chicago. Intern.
Zoo Ybk. 12:218-220, 1972.
Meritt,
D.A.: Edentate diets. II. Two-toed sloths. Lab. An. Sci. 23:543-543, 1973.
Meritt, D.A.: A further note on the immobilisation of sloths. Intern.
Zoo Ybk. 14:160-61, 1974.
Meritt,
D.A.: Edentates. The Ark (Lincoln Park Chicago) 2:2-14, 1975.
Meritt, D.A.: A simplified technique for diagnosing parturition in Hoffmann's
sloth. Intern. Zoo Ybk. 16:152-153, 1976.
Meritt,
D.A.: The natural history, behavior, nutrition, physiology, reproduction,
development and management of Hoffmann's sloth, Choloepus hoffmanni
(Peters). M.A. Thesis, Northeastern Illinois University, 124 pp., 1977.
(Kindly supplied by author).
Meritt, D.A.: The two-toed Hoffmann's sloth, Choloepus hoffmanni
Peters. In, The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas.
G.G. Montgomery, ed. Smithsonian Institute Press, Washington, D.C., pp.
333-341, 1985.
Moser,
H.G. and Benirschke, K.: The fetal zone of the adrenal gland in the nine?banded
armadillo, Dasypus novemcinctus. Anat. Rec. 143:47?60, 1962.
Murata,
K. and Masuda, R.: Gender determination of the Linné's two-toed
sloth (Choloepus didactylus) using SRY amplified from hair. J.
Vet. Med. Sci. 58:1157-1159, 1996.
Nowak,
R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press,
Baltimore, 1999.
Seymour,
C., Peralta, P.H. and Montgomery, G.G.: Viruses isolated from Panamanian
sloths. Amer. J. Trop. Med. Hyg. 32:1435-1444, 1983.
Soma,
H.: In chase of trophoblast: A comparison between human and other mammalian
placentas. J. Tokyo Med. Coll. 34:873-883, 1976 (in Japanese).
Sonta
S.-i., Hayata, I., Sasaki, M. and Kondo, N.: The karyotypes and sex-determining
mechanism in the two-toed sloth, Choloepus didactylus. Chromos.
Inf. Serv. 28:15-17, 1980.
Veselovsky,
Z.: A contribution to the knowledge of the reproduction and growth of
the two-toed sloth Choloepus didactylus at Prague Zoo. Int. Zoo
Ybk. 6:147-153, 1966.
Wetzel,
R.M.: Systematics, distribution, ecology, and conservation of South American
edentates. Pp. 345-375, in "Mammalian Biology in South America. Special
Publication Series of the Pymatuning Laboratory of Ecology, University
of Pittsburg, Vol. 6, 1982.
|