| |
16) Physiological data
No publications are known to me.
17)
Other resources
Fibroblast strains are available through CRES at the San Diego Zoo.
Pregnancy and birth of a common chimpanzee following artificial insemination
with post mortem frozen epididymal sperm has been reported by Kusunoki
et al. (2001).
References
CRES at: http://www.sandiegozoo.org/conservation/cres_home.html
Benirschke, K. and Kumamoto, A.T.: Paternity diagnosis in pygmy chimpanzees.
Intern. Zoo Ybk. 23:220-223, 1985.
Benirschke,
K., Bogart, M.H., McClure, H.K. and Nelson-Rees, W.A.: Fluorescence of
the trisomic chimpanzee chromosomes. J. Med. Primatol. 3:311-314, 1974.
Benirschke,
K., Bogart, M.H. and Adams, F.: The status of the pygmy chimpanzee. Intern.
Zoo Ybk. 20:71-76, 1980.
Bourne,
G.H., ed. The Chimpanzee, Vol. 1: Anatomy, Behavior, and Diseases of Chimpanzees.
University Park Press, Baltimore, 1969. (and Karger, Basel).
Bogart,
M.H. and Benirschke, K.: Chromosomal analysis of the pygmy chimpanzee
(Pan paniscus) with a comparison to man. Folia Primatol. 27:60-67,
1977a.
Bogart,
M.H. and Benirschke, K.: Q-band polymorphism in a family of pygmy chimpanzee
(Pan paniscus). J. Med. Primatol. 6:172-175, 1977b.
Bowman,
M.E., Lopata, A., Jaffe, R.B., Golos, T.G., Wickings, J. and Smith, R.:
Corticotropin-releasing hormone-binding protein in primates. Amer. J.
Primatol. 53:123-130, 2001.
Chou, H.-H., Takematsu, H., Diaz, S., Iber, J., Nickerson, E., Wright,
K.L., Muchmore, E.A., Nelson, D.L., Warren, S.T. and Varki, A.: A mutation
in human CMP-sialic acid hydroxylase occurred after the Homo-Pan divergence.
Proc. Natl. Acad. Sci. 95:11751-11756, 1998.
Coolidge,
H.J.: Pan paniscus (Pygmy chimpanzee) from south of the Congo River.
Amer. J. Phys. Anthropol. 8:1-57, 1933.
Czekala,
N.M., Hodges, J.K. and Lasley, B.L.: Pregnancy monitoring in diverse primate
species by estrogen and bioactive luteinizing hormone determinations in
small volumes of urine. J. Med. Primatol. 10:1-15, 1981.
de
Waal, F. and Lanting, F.: Bonobo. The Forgotten Ape. University of California
Press, Berkeley, 1997.
Jones,
S., Martin, R. and Pilbeam, D., eds.: The Cambridge Encyclopedia of Human
Evolution. Cambridge University Press, Cambridge 1995.
Jurke,
M.H., Hagey, L.R., Jurke, S. and Czekala, N.M.: Monitoring hormones in
urine and feces of captive bonobos (Pan paniscus). Primates 41:311-319,
2000.
Khudr,
G., Benirschke, K. and Sedgwick, C.J.: Man and Pan paniscus: A karyologic
comparison. J. Human Evol. 2:323-331,1973.
Kusunoki,
H., Daimaru, H., Minami, S., Nishimoto, S., Yamane, K.-I. and Fukumoto,
Y.: Birth of a chimpanzee (Pan troglodytes) after artificial insemination
with cryopreserved epididymal spermatozoa collected postmortem. Zoo Biol.
20:135-143, 2001.
Lasley,
B.L., Kennedy, J.F., Robinson, P.T., Bogart, M.H. and Benirschke, K.:
A study of reproductive failure in a pygmy chimpanzee (Pan paniscus).
Zool. Garten 47:289-295, 1977.
Lear, T.L., Houck, M.L., Zhang, Y.W., Debnar, L.A., Sutherland-Smith,
M.R., Young, L., Jones, K.L., and Benirschke, K.: Trisomy 17 in a bonobo
(Pan paniscus) and deletion of 3q in a lowland gorilla (Gorilla
gorilla gorilla): comparison with human trisomy 18 and human deletion
4q syndrome. Cytogenet. Cell Genet. 95:228-233, 2001).
Puschmann,
W.: Zootierhaltung. Vol. 2, Säugetiere. VEB Deutscher Landwirtschaftsverlag,
Berlin, 1989.
Scott,
G.B.D.: Comparative Primate Pathology. Oxford University Press, Oxford,
1992.
Smith, A., Skilling, D.E., Ensley, P.K., Benirschke, K. and Lester, T.L.:
Calicivirus isolation and persistence in a pygmy chimpanzee (Pan paniscus).
Science 221:79-81, 1983.
Additional consideration of the placentation
in the common chimpanzee,
Pan troglodytes
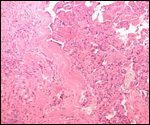
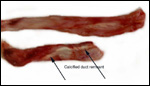
Several placentas have become available to us that differ only in minor
detail from that of the bonobo. Thus, the term placenta of one such animal
was 300 g, had an eccentric umbilical cord insertion, but the cord was
only 26 cm long. It had a chalky streak, which was interpreted as being
a calcified vitelline duct remnant. The remainder was normal.
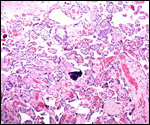
Another placenta had typical "X-cell" cysts (extravillous trophoblast).
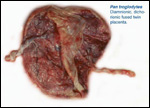
One additional placenta was of fraternal twins (male/female). It was kindly
made available by Mrs. Jo Fritz (Arizona). It was from a premature delivery
and was an example of a fused diamnionic dichorionic twin placenta, weighing
230 g combined. One cord was 15 cm, the other 49 cm long and one had a
right, the other a left twist; one was eccentric, the other was marginal
in its insertion. One had funisitis, a common cause of premature birth
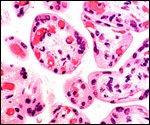
in human pregnancies. An unexplained feature of this specimen was the
large number of hemosiderin-laden macrophages in the villi of twin 2.
|