 |
|
(Clicking
on the thumbnail images below will launch a new window and a larger
version of the thumbnail.)
|
| Last updated: February 25, 2005. |
Bos (Bibos) javanicus
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
The Banteng is now classified as an endangered species, having been reduced in numbers significantly in most of its former habitat. This large bovid clusters with gaur in a detailed, modern molecular study by Buntjer et al. (2002). They are large, brown animals weighing up to 900 kg. Males are much darker and have longer horns. The name "banteng" is said to be of Malayan origin (Gotch, 1979). There is a good breeding colony at the San Diego Zoo from which these specimens have come. The longevity of bantengs is 26 years 7 months according to Jones (1993). Nowak (1999) reported that there are now 1.5 million domestic banteng ("Bali cattle") used for domestic work and meat. Feral Banteng also exist in Australia, originating from 20 imported animals in 1849. Numerous studies have examined the hemoglobin and other proteins in these Australian Bali cattle and these were compared them with other breeds (e.g., Bachman et al., 1978; Bell et al., 1981; Namikawa et al., 1983; Bell et al., 1990).
 |
Adult male and female bantengs at the San Diego Wild Animal Park. |
 |
Juvenile bantengs at the San Diego Wild Animal Park. |
Banteng pregnancy lasts 9.5-10 months and it usually produces a single young. Twins are not uncommon, however. The estrous cycle lasts 3-4 weeks and sexual maturity is reached at 2 years. Newborns weigh around 30 kg (Puschmann, 1989). It has recently been possible to obtain calves by artificial insemination (Johnston et al., 2002).
3) Implantation
No data have been reported on early implantation of banteng placentas. It is likely to be similar as in cattle since hybridization and embryo transfer are possible. Both uterine horns are employed in placentation.
4)
General Characterization of the Placenta
Two full term placentas were available from healthy, surviving female
newborns. They had a cotyledonary placenta with 25 and 53 flat cotyledons,
respectively. They measured 25 x 12 to 5 mm, and 14 x 54 mm in width,
respectively, and were 1-1.5 cm in thickness. The cotyledons were arranged
in four rows, much like those in cattle. One (probably incomplete) placenta
weighed 1,250 g, the other was 3,000 g. They had central cord insertions.
In calculating the cotyledonary surface area as suggested by Wild (1964),
it comes to approximately 950 cm2 in the first placenta and 1,100 cm2
in the second, considerably less than for values of neonatal calves (+/-
4500 cm2). Perhaps the extremely long, finely divided villi compensate
for the relatively small surface area of cotyledons. Apparently the banteng
placenta occupies both uterine horns. The lesser horn has much smaller
cotyledonary sizes.
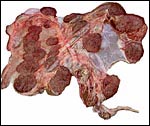 |
Placenta (lacking the portion of the lesser horn) of term banteng with 25 large cotyledons. |
 |
Single cotyledon after fixation. |
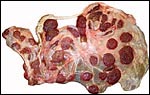 |
Second placenta with a minor portion located in the lesser uterine horn at left. |
These placentas are typical of most ungulate placentas in their histological appearance. They are cotyledonary, with major stem villi and fine branching of villous structures. The villi were covered by a single layer of trophoblast among which numerous binucleate cells were found. These were considerably larger than the other trophoblastic cells. As in most other ungulate placentas described here, the binucleate cells are presumed to be responsible for placental lactogen production. These aspects are further described in the deer placenta and that of other ungulates. Beneath the chorionic surface, the trophoblast is more cylindrical and has a paler cytoplasm. It is also frequently pigmented in a manner that is also described in detail in the descriptions of other ungulate placentas. The pigment does not stain for iron and its nature, as in other ungulates, is still unknown. While this region (and pigment) has been referred to as the "haemophagous organ" (speculated to contribute with iron transfer to the fetus from focal maternal hemorrhages), this is highly speculative in my opinion. Fresh blood is virtually never found in these locations.
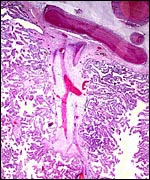 |
Stem villus with branching villi of this delivered placenta. |
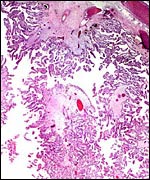 |
Surface of placenta with chorion and fetal vessels at top. |
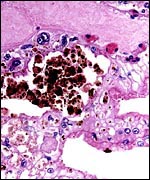 |
Beneath the chorionic surface is a layer of tall trophoblast with intense (iron-negative) yellow-brown pigment. |
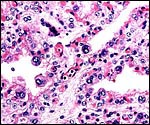 |
Higher magnification of villi with at least three large, typical binucleate cells. |
The umbilical cords measured 10 cm and 22 cm in length, had four vessels, an allantoic duct and no twists. Numerous small vessels were also present in sections of the umbilical cord. No verrucous projections were found on the surface of these largely allantoic portions of the cords.
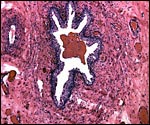 |
Allantoic duct in the center of the umbilical cord, accompanied by numerous small blood vessels. The red content of the duct is similar to hippomanes. |
There are no reports.
8)
Extraplacental membranes
There is a large allantoic sac with finely distributed vasculature. A
small amount (2 x 1 cm) of light brown hippomanes was present within the
second placenta.
 |
Hippomanes enclosed in allantoic membrane. |
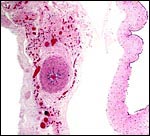 |
Membranes, with amnion (avascular) at right and allantois (vascularized) at left. Allantoic epithelium is degenerated. |
I assume that, like other ungulates, there is no trophoblast invasion of the uterus.
10)
Endometrium
None was available in these two delivered placentas.
11)
Various features
None.
12) Endocrinology
I know of no reports on endocrine studies in this species.
13)
Genetics
Banteng have 60 acrocentric chromosomes, similar to cattle, except for
minor rearrangements seen on banding studies. A very detailed analysis
of bovid chromosomes was undertaken by Gallagher et al. (1999). It showed,
i.a., that the Y-chromosome of banteng is metacentric, even though the "original" Y-chromosome was inferred to be an acrocentric element.
Mayr et al. (1985) defined the centromeres and NORs in various bovidae,
including the banteng. Pathak & Kieffer (1979) also studied centromeric
heterochromatin and found it in all autosomes but little was present in
the X-chromosomes. The Y-chromosome had an appreciable amount of heterochromatin.
Hybrid bulls had arrest of spermatogenesis at pachytene. Potter & Upton (1979) and Potter et al. (1979) studied banding patterns of the
Y-chromosome and were able to differentiate among different bovid species.
Hybrids with gaur (Bos gaurus) are said to be fertile (Nowak, 1999).
On the other hand, Steklenev & Elistratova (1992) reported that male
hybrids between banteng and domestic cow are sterile, while females are
fertile. Gray (1972) also reported on hybrids with domestic cattle.
 |
Karyotype of male banteng. Note the small metacentric Y-chromosome. |
 |
Banteng calf from embryo transfer into Simmental cow (Munich Zoo, 1983). |
The San Diego Zoo exhibits now the first successfully cloned male banteng. The source of cells was a male that died in a battle between bulls; it had been born in 1974 and the fibroblasts had been kept in the “Frozen Zoo” of the Society. With the help of a Biotech Company, an enucleated ovum from a Simmental cow was used to place the fibroblast nucleus and a normal offspring resulted that is shown next. Its undescended testes underwent orchiopexy at about one year of age and the young bull (now 2 years old in 2005) is placed with females. It is hoped that its rare genotype will be ‘reinjected’ into the banteng herd.
 |
Neonatal cloned banteng bull. |
 |
Cloned banteng bull at 2 years of age. |
14)
Immunology
I am not aware of any immunological studies.
15)
Pathological features
Griner (1983) examined many banteng autopsies from the colony at the San
Diego Zoo. He found omphalitis once in a newborn and many cases having
died from malignant catarrhal fever (MCF) virus infection. Additional
details on MCF may be found in the publication by Hatkin (1980). Similar
outbreaks of MCF occurred at the Munich Zoo (Hanichen et al., 1998). Infection
with Sarcocystis spp. was described by Odening et al. (1995).
Wild (1964) measured the surface of chorion and, separately, that of the
cotyledons in domestic cow placentas in order to ascertain a possible
relation to neonatal outcome. Whether this applies to banteng (whose cotyledonary
surface is one fourth that of cattle) is unknown but will be of interest
when banteng placentas from clones into cows become available.
16)
Physiologic data
I know of no physiologic studies in this species, other than the protein
electrophoretic reports mentioned at the beginning.
17)
Other resources
Strains of fibroblast cultures from several banteng at the San Diego Zoo
may be obtained by contacting Dr. Oliver Ryder at oryder@ucsd.edu.
18)
Other remarks - What additional Information is needed?
Early implantational stages, and implanted placentas as well will be of
interest in the future. The larger, flat cotyledons of this species differ
much from the cotyledons of cattle with which they should be compared
in better detail in the future also.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society
of San Diego. I appreciate also very much the help of the pathologists
at the San Diego Zoo.
References
Bachmann, A.W., Campbell, R.S. and Yellowlees, D.: Haemoglobin in cattle
and buffalo. Haemoglobin types of Bos taurus, Bos indicus,
Bos banteng and Bubalis bubalis in northern Australia. Australian
J. Exp. Biol. Med. Sci. 56:523-529, 1978.
Bell, K., Hopper, K.E. and McKenzie, H.A.: Bovine alpha-lactalbumin C and alpha S1-, beta- and kappa-caseins of Bali (Banteng) cattle Bos [Bibos] javanicus. Australian J. Biol. Sci. 34:149-159, 1981.
Bell, K., McKenzie, H.A. and Shaw, D.C.: Hemoglobin, serum albumin and transferring variants of Bali (Banteng) cattle, Bos [Bibos] javanicus. Comp. Biochem. Physiol. B 95:825-832, 1990.
Buntjer, J.B., Otsen, M., Nijman, I.J., Kuiper, M.T. and Lenstra, J.A.: Phylogeny of bovine species based on AFLP fingerprinting. Heredity 88:46-51, 2002.
Gallagher, D.S., Davis, S.K., DeDonato, M., Burzlaff, J.D., Womack, J.E., Taylor, J.F. and Kumamoto, A.T.: A molecular cytogenetic analysis of the tribe Bovini (Artiodactyla: Bovidae: Bovinae) with an emphasis on sex chromosome morphology and NOR distribution. Chromosome Res. 7:481-492, 1999.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Gray,
A.P.: Mammalian Hybrids. A Check-list with Bibliography. 2nd edition.
Commonwealth Agricultural Bureaux Farnham Royal, Slough, England, 1972.
Griner, L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego, California, 1983.
Hanichen, T., Reid, H.W., Wiesner, H. and Hermanns, W.: Malignant catarrhal fever in zoo ruminants. Tieraerztl. Prax. Ausg. G. Grosstiere Nutztiere. 26:294-300, 1998 (in German).
Hatkin, J.: Endemic malignant catarrhal fever at the San Diego wild animal park. J. Wildl. Dis. 16:439-443, 1980.
Johnston, S.D., McGowan, M.R. and Blyde, D.: Birth of a Banteng (Bos javanicus) calf at Western Plains Zoo after fixed time artificial insemination. Australian Vet. J. 80:94-95, 2002.
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Mayr, B., Schweizer, D., Mendelak, M., Krutzler, J., Schleger, W., Kalat, M. and Auer, H.: Levels of conservation and variation of heterochromatin and nucleolus organizers in the Bovidae. Can. J. Genet. Cytol. 27:665-682, 1985.
Namikawa, T., Takenaka, O. and Takahashi, K.: Hemoglobin Bali (bovine): beta A 18(Bl)Lys leads to His: one of the "missing links" between beta A and beta B of domestic cattle exists in the Bali cattle (Bovinae, Bos banteng). Biochem. Genet. 21:787-796, 1983.
Nowak, R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press, Baltimore, 1999.
Odening, K., Wesemeier, H.H., Walter, G. and Bockhardt, I.: On the morphological diagnostics and host specificity of the Sarcocystis species of some domesticated and wild bovini (cattle, Banteng and bison). Appl. Parasitol. 36:161-178, 1995.
Pathak, S. and Kieffer, N.M.: Sterility in hybrid cattle. I. Distribution of constitutive heterochromatin and nucleolus organizer regions in somatic and meiotic chromosomes. Cytogenet. Cell Genet. 24:42-52, 1979.
Potter, W.L. and Upton, P.C.: Y chromosome morphology of cattle. Australian Vet. J. 55:539-541, 1979.
Potter, W.L., Upton, P.C., Cooper, J. and Blackshaw, A.W.: C- and G-banding patterns and chromosomal morphology of some breeds of Australian cattle. Australian Vet. J. 55:560-567, 1979.
Puschmann, W.: Zootierhaltung. Vol. 2, Säugetiere. VEB Deutscher Landwirtschaftsverlag Berlin, 1989.
Steklenev, E.P. and Elistratova, T.M.: The characteristics of reproductive capacity of hybrids of banteng (Bos [Bibos] javanicus d'Alton) with the domestic cow (Bos [Bos] primigenius taurus). Tsitol. Genet. 26:45-57, 1992 (in Russian).
Wiesner, H., Lempeter, W.W. and Rietschel, W.: Erfahrungen beim unblutigen Embryotransfer vom Banteng auf Hausrinder. Verhandl. 26. Intern. Sympos. Erkrankungen der Zootiere. Brno, 1984. Akademie-Verlag, Berlin, pp.99-102, 1984.
Wild, A.: Untersuchungen über den Aufbau der Placenta fetalis des Rindes und ihre Auswirkungen auf die Gesundheit des Kalbes. Zbl. Veterinärmedizin "B". 11:60-89, 1964.