| |
References
Adamoli, V.C., Cetica, P.D., Merani, M.S. and Solari, A.J.: Comparative
morphologic placental types in Dasypodidae (Chaetophractus villosus,
Cabassous chacoensis, Tolypeutes matacus and Dasypus hybridus).
Biocell 25:17-22, 2001.
Affanni,
J.M., Samartino, L.G., Casanave, E.B. and Dezi, R.: Absence of apnea in
armadillos covered by soil. Resp. Physiol. 67:239-245, 1987.
Amborski,
R.L., LoPiccolo, G. and Amborski, G.F.: Development of an established
cell line derived from Dasypus novemcinctus, a laboratory animal
susceptible to infection by Mycobacterium leprae. Experientia 30:546-548,
1974.
Anderson,
J.M. and Benirschke, K.: Fetal circulations in the placenta of Dasypus
novemcinctus, Linn. and their significance in tissue transplantation.
Transplantation 1:306, 1963 .
Arruda,
O.S. de and Opromolla, D.V.A. Manutença de tatus em cativero (Mammalia,
Edentata). Maintenance of armadillos in Captivity (Mammalia, Edentata).
Revista Biotérios 1:57-62, 1981.
Barreto,
M., Barreto, P. and D'Allessandro, A.: Colombian armadillos: Stomach content
and infection with Trypanosoma cruzi. J. Mammal. 66:188-193, 1985.
Beath,
M.M. Benirschke, K.. and Brownhill, L.E.: The chromosomes of the nine-banded
armadillo, Dasypus novemvinctus). Chromosoma 13:27, 1962.
Benirschke,
K., Sullivan, M.M. and Marin-Padilla, M.: Size and number of umbilical
vessels. A study of multiple pregnancy in man and armadillo. Obstet. Gynecol.
24:819-834, 1964.
Billingham,
R.E. and Neaves, W.: Exchanges of skin-grafting among monozygotic quadruplets
in armadillos. J. Exper. Zool. 213:257-260, 1980.
Brinck-Johnsen,
T., Benirschke, K. and Brinck-Johnsen, K.: Hormonal steroids in armadillo,
Dasypus novemcinctus. 1. Oestriol in pregnancy and its in vitro
biosynthesis by the placenta. Acta Endocrinol. 56:675-690, 1967.
Buchanan,
G.D., Enders, A.C. and Talmage, R.V.: Implantation in armadillos ovariectomized
during the period of delayed implantation. J. Endocrinol. 14:121, 1956.
Buchanan, G.D.: Variation in litter size of nine-banded armadillos. J.
Mammal. 38:529, 1957.
Buchanan,
G.D.: The current range of the armadillo Dasypus novemcinctus mexicanus
in the United States. Texas J. Sci. 10:349-351, 1958.
Buchanan,
G.D.: The presence of two conceptuses in the uterus of a nine-banded armadillo.
J. Reprod. Fert. 13:329-331, 1967.
Cell
lines from: http://www.sandiegozoo.org/conservation/cres_home.html.
Please direct your inquiries to Dr. Oliver Ryder (oryder@ucsd.edu).
Cuba-Caparó,
A.: Some hematologic and temperature determinations in the 7-banded armadillo
(Dasypus hybridus) Lab. Anim. Sci. 26:450-455, 1976.
Datta,
M.: Study of meiotic chromosomes of the nine-banded armadillo, Dasypus
novemcinctus. The Nucleus 12:69, 1969.
Ebaugh,
F.G. and Benson, M.A.: Armadillo hemoglobin characteristics and red cell
survival. J. Cell. Comp. Physiol. 64:183-192, 1964.
Enders,
A.C.: The structure of the armadillo blastocyst. J. Anat. 96:39, 1962.
Enders, A.C.: Development and structure of the villous haemochorial placenta
of the nine-banded armadillo (Dasypus novemcinctus). J. Anat. 94:34-45,
1960.
Enders, A.C.: The reproductive cycle of the nine-banded armadillo (Dasypus
novemcinctus). In, Comparative Biology of Reproduction in Mammals,
I.W. Rowlands, ed. Symposia of the Zoological Society of London, # 15.
Academic Press London, 1966. pp. 295-310.
Enders,
A.C.: Electron microscopic observations on the villous hemochorial placenta
of the nine-banded armadillo (Dasypus novemcinctus). J. Anat. 94:205,
1960.
Enders, A.C.: Implantation in the nine-banded armadillo: How does a single
blastocyst form four embryos? Placenta 23:71-85, 2002.
Enders,
A.C. and Buchanan, G.D.: The reproductive tract of the female nine-banded
armadillo. Texas Rep. Biol. Med. 17:323, 1959.
Enders, A.C. and Welch, A.O.: Structural interactions of trophoblast and
uterus during hemochorial placenta formation. J. Exp. Zool. 266:578-587,
1993.
Frechkop,
S. and Yepes, J.: Études systématique et zoogéographique
des dasypodidés conservés a l'institut. Bull. Inst. Roy.
Sci. Nat. Belg. 25:1-56, 1949.
Galbreath,
G.J.: The evolution of monozygotic polyembryony in Dasypus. In,
The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas. G.G.
Montgomery, ed. Smithsonian Institution Press, Washington, 1985. pp.243-246.
Gray,
J.E.: Handlist of Edentates, Thick-skinned and ruminant mammals in the
British Museum. Section 2 - Loricata. British Mus., London, pp.11-25,
1873.
Hamlett,
G.W.D.: The reproductive cycle in the armadillo. Z. wiss. Zool. A. 141:143,
1932.
Hamlett,
G.W.D.: Identity of Dasypus septemcinctus Linnaeus with notes on some
related species. J. Mammal. 20:328-336, 1939.
Humphrey,
S.R.: Zoogeography of the nine-banded armadillo (Dasypus novemcinctus).
BioSci. 24:457-462, 1974.
Labhsetwar,
A.P. and Enders, A.C.: Progesterone in the corpus luteum and placenta
of the armadillo, Dasypus novemcinctus. J. Reprod. Fert. 16:381-387, 1968.
Lasley,
B.L., Czekala, N.M., Nakakura, K.C., Amara, S.G. and Benirschke, K.: Armadillos
for studies on delayed implantation, quadruplets, uterus simplex, and
fetal adrenal physiology. In, Animal Models for Research on Contraception
and Fertility. N.J. Alexander, ed. Harper and Row, Hagerstown, MD 1979.
pp. 447-451.
Layne,
J.N. and Glover, D.: Home range of the armadillo in Florida. J. Mammal.
58:411-413, 1977.
Loughry,
W.J., Prodöhl, P.A., McDonough, C.M. and Avise, J.C.: Polyembryony
in armadillos. Amer. Scientist 86:274-279, 1998.
Marin-Padilla,
M. and Benirschke, K.: Thalidomide induced alterations in blastocyst and
placenta of the armadillo, Dasypus novemcinctus mexicanus, including
a choriocarcinoma. Amer. J. Pathol. 43:999-1016, 1963.
Meritt,
D.A.: Edentate diets. I. Armadillos. Lab. Anim. Sci. 23:540-542, 1973.
Mossman,
H.W.: Vertebrate Fetal Membranes. MacMillan Press, Houndmills, 1987.
Newman, H.H. and Patterson, J.T.: The development of the nine-banded armadillo
from the primitive streak stage to birth, with especial reference to the
question of specific polyembryony. J. Morphol. 21:359, 1910.
Newman,
H.H. and Patterson, J.T.: The limits of hereditary control in armadillo
quadruplets: A study of blastogenic variation. J. Morphol. 22:855-926,
1911.
Parkes,
A.S.: Marshall's Physiology of Reproduction. 3rd ed. Vol. I. p. 237. Longmans,
Green & Co., London, 1956.
Peppler,
R.D. and Stone, S.C.: Plasma progesterone level during delayed implantation,
gestation and postpartum period in the armadillo. Lab. Anim. Sci. 30:188-191,
1980.
Peppler,
R.D. and Stone, S.C.: Annual pattern in plasma testosterone in the male
armadillo, Dasypus novemcinctus. Anim. Reprod. Sci. 4:49-53, 1981.
Peppler,
R.D. and Canale, J.: Quantitative investigation of the annual pattern
of follicular development in the nine-banded armadillo (Dasypus novemcinctus).
J. Reprod. Fertil. 59:193-197, 1980.
Phillips,
J.A., Harlow, H.J., McArthur, N.H. and Ralph, C.L.: Epithalamus of the
nine-banded armadillo, Dasypus novemcinctus. Comp. Biochem. Physiol.
85A:477-481, 1986.
Prodöhl,
P.A., Loughry, W.J., McDonough, C.M., Nelson, W.S. and Avise, J.C.: Molecular
documentation of polyembryony and the micro-spatial dispersion of clonal
sibships in the nine-banded armadillo, Dasypus novemcinctus. Proc.
R. Soc. Lond. B - Biol. Sci. 263:1643-1649, 1996.
Ramsey,
E.M.: The Placenta. Human and Animal. Praeger, NY 1982.
Rideout,
B.A., Gause, G.E., Benirschke, K. and Lasley, B.L.: Stress-induced adrenal
changes and their relation to reproductive failure in captive nine-banded
armadillos (Dasypus novemcinctus). Zoo Biol. 4:129-137, 1985.
Scholander,
P.F., Irving, L. and Grinell, S.W.: Respiration of armadillo with possible
implication as to its burrowing. J. Cell Comp. Physiol. 21:53-63, 1943.
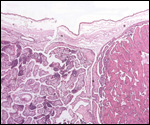
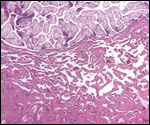
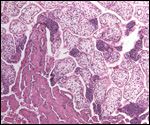
Smith, M.D. and Procop, G.W.: Typical histologic features of Tunga
penetrans in skin biopsies. Arch. Pathol. Lab. Med. 126:714-716, 2002.
Stockard, C.R.: A probable explanation of polyembryony in the armadillo.
Amer. Naturalist 55:62-68, 1921.
Storrs,
E.E.: Animal Model: Experimental lepromatous leprosy in nine-banded armadillos
(Dasypus novemcinctus). Amer. J. Pathol. 92:813-816, 1978.
Storrs,
E.E., Burchfield, H.P. and Rees, R.J.W.: Superdelayed parturition in armadillos:
a new mammalian survival strategy. Lepr. Rev. 59:11-15, 1988.
Storrs,
E.E. and Williams, R.J.: A study of monozygous quadruplet armadillos in
relation to mammalian inheritance. Proc. Natl. Acad. Sci. 60:910-914,
1968.
Storrs,
E.E., Walsh, G.P. and Burchfield, H.P.: Development of leprosy in another
species of armadillo Dasypus hybridus (L.): Genetic and immunologic
implications. J. Trop. Med. Hyg. 78:216-218, 1975.
Strauss,
F.: Probleme der Ovo-Implantation bei den Säugetieren. Z. Säugetierk.
46:65-79, 1981.
Strozier,
L.M., Blair, C.B. and Evans, B.H.: Armadillos: Basic profiles. I. Serum
chemistry values. Lab. Anim. Sci. 21:399-400, 1971.
Wetzel,
R.M. and Mondolfi, E.: The subgenera and species of long-nosed armadillos,
genus Dayspus L. In, Eisenberg, J.F. ed. Vertebrate Ecology in the Northern
Neotropics. pp. 43-63, Smithsonian Inst. Press, Washington, DC, 1979.
|