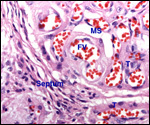
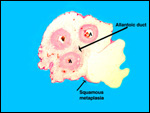
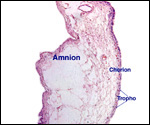
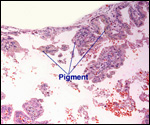
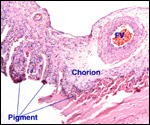
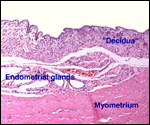
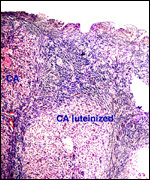
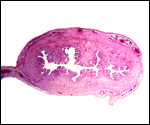
|
|
(Clicking
on the thumbnail images below will launch a new window and a larger
version of the thumbnail.)
|
| Last updated: July 23, 2005. |
Orycteropus afer
Order: Tubulidentata
Family: Orycteropodidae
1) General zoological data
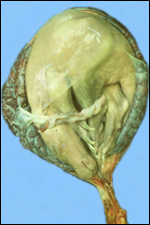
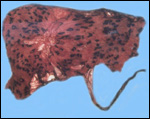
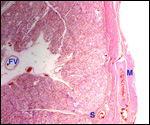
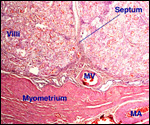
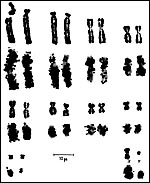
This single species of this mammalian Order lives in the sub-Saharan countries of Africa and is primarily nocturnal. The fossil records show that it extended once to the Mediterranean region. Its precise taxonomic relationship to other mammals has been a problem for classification. Perhaps this, as well as its unusual architecture (teeth, skin, tongue, etc.), is reason enough for a separate Order status among the mammals. Current genetic studies to clarify its taxonomic position are discussed under genetics below. Adults weigh 40-100 kg, most are around 50-70 kg (Nowak, 1999). The teeth lack enamel and grow continuously. Aardvarks ("earth-pigs") burrow deep holes and consume large quantities of ants and termites. They have their single young in deep tunnels, are solitary, and have a very characteristic appearance. They are allegedly good swimmers. Aardvarks have poor eyesight but can hear and smell exquisitely well. They are solitary, males are slightly larger than females. They have a very long tongue and extremely tough skin. Externally, it is hard to determine their sex. Aardvarks do well in captivity. They can be trained and have repeatedly reproduced in different zoological gardens (see Dulaney, 1987). An interesting finding of aardvarks is that they do not have a clavicle.
2)
General gestational data
The gestation is 7-9 months long, according to Hayssen et al. (1993).
Twins are very uncommon. Newborns weigh approximately 2 kg. In captive
births, the newborns are being "marked" frequently by the scent
gland at the mother's tail (Dulaney, 1987). Sexual maturity is reached
at 2 years of age.