 |
| Last updated: October 3, 2010. |
Three-banded armadillo
Tolypeutes matacus
Order: Xenarthra (Edentata)
Family: Dasypodidae
1) General Zoological Data
The name Tolypeutes derives from “tolupē” (Gr.) = ’ wool made into a ball, a ball wound up’ (Gotch, 1979). The scientist having the most experience with this species is Dennis Meritt of Chicago who has kept these animals at Lincoln Park Zoo and studied their biology the most diligently. 18 offspring were produced at that zoo. The parents of our specimen had come from Paraguay, but the animals also live in Bolivia, Argentina and Brazil (Eisentraut, 1933). A breeding colony was established at the San Diego Zoo in the 1990s. This species, often referred to, or being identical with Tolypeutes tricinctus, produces normally a single embryo after a gestation of 120 days (Meritt, 1971). Adults of this normally insectivorous animal weigh 1-1.59 kg and may live up to 17 years in captivity (Nowak, 1999). As is seen in Figure 1, the three-banded armadillo often has an irregular band structure (2-4 vs. 3), as is a common feature also in other Dasypodidae (Nowak, 1999). Thenius & Hofer (1960) believe that the time of origin of this species is poorly understood, lacking fossils. Thenius (1969) provides further evidence of the relationship to other mammalian forms and gave data on bone and scute structure. A detailed review of the relationships of these animals was presented by Wetzel et al. (2007). This South American armadillo is the only species that is able to roll up into a ball (second picture). Specifics about the maintenance of Xenarthra in captivity are discussed by Puschmann (2004).
2) General gestational data
This male newborn three-banded armadillo was delivered by Cesarean section because of presumed dystocia. It weighed 101.2 g at birth but died two days later when its weight was 97.6 g. The cause of death was massive pulmonary hemorrhage. See also Hayssen et al. (1993) for an extensive survey of mammalian biology.
 |
Gross appearance of neonatal three-banded armadillo. |
 |
The only armadillo species that can roll up into a ball and was named for that ability. |
3) Implantation
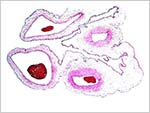
No implanted placenta has yet been observed. From the appearance of this delivered placenta, however, one would expect that it is superficially implanted on a deciduas basalis. The delivered placenta weighed 28 g and measured 9.5 X 7 x 0.4 cm and grossly appeared to be villous.
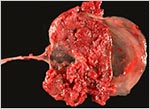 |
Gross appearance of the fragmented placenta with cord extending to the left. |
4) General characteristics of placenta
This is a villous placenta with remarkable histological similarity to the human placenta. A single layer of trophoblast covers the villi. The villi contain Hofbauer cells (macrophages) and capillaries which are endowed with a simple endothelium. The trophoblast peripherally has large nuclei but true syncytial cells have not been observed. The floor of the placenta has a layer of fibrin, very much like Nitabuch’s fibrin layer. Fetal vessels are carried in the chorionic membrane which lies beneath a simple, avascular amnionic membrane. Numerous clusters of extravillous trophoblast (so-called “X-cells”) are present throughout the villous tissue. These often contain some fibrinoid.
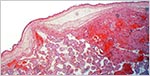 |
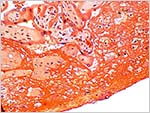
Margin of the placenta with minimal infarction at top right. It shows the membranes very well. |
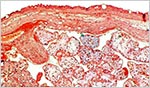 |
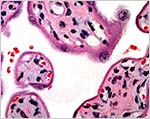
Surface of the term placenta showing the island of extravillous trophoblast at left. |
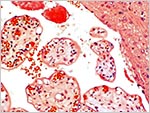 |
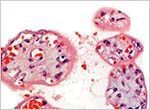
Mature villi and extratrophoblastic island at right. |
 |
Base of the placenta with Nitabuch fibrin layer. |
 |
Trophoblastic surface of villi. |
 |
Villous surfaces. |
5) Details of placental barrier
A single layer of trophoblast covers the villi and the surface appears to have microvilli. It will be necessary, however, to obtain EM pictures of this structure before villous cytotrophoblast can be ruled out as possible precursor of the outer layer.
6) Umbilical cord
The cord attached to the placenta was 3.9 x 0.4 cm and had 4 blood vessels, two arteries and two veins. No ducts were identified, thus there is no allantoic sac. It appeared that the cord was relatively flat in its composition. Additional cord was present on the neonate that is seen in Figure 1.
 |
Gross appearance of the fragmented placenta with cord extending to the left. |
7) Uteroplacental circulation
This has not been studied.
8) Extraplacental membranes
The membranes of this placenta consist of only amnion and chorion. No extravillous trophoblast surrounds the membranes and there is no deciduas capsularis. The chorion of this placenta, however, carried a few fetal blood vessels.
9) Trophoblast external to barrier
Extravillous trophoblast infiltrates the base of the placenta which has a distinct Nitabuch’s fibrin layer.
10) Endometrium
At the base of some sections of the placenta small fragments of deciduas basalis are attached. The uterus is simple and pear-shaped (Cetica et al. (2005).
11) Various features
The number of “bands” in the carapace varies slightly, from 2 to 4.
12) Endocrinology
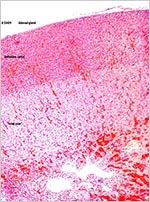
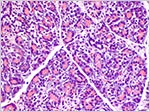
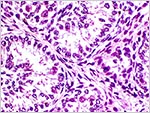
The adrenal gland is distinctly unusual. It has a broad definitive cortex and a nearly equally wide fetal cortex (see next figure). Fetal zones of the placenta are rarely found in non-mammalian species, but the nine-banded armadillo is another exception (Brinck-Johnsen et al., 1967, 1970). In human fetuses it produces DEA that is aromatized to estriol that is present in abundance in pregnant women. The testis and epididymis were well developed. Testicular tubules contained germ cells but the interstitial Leydig cells were not stimulated; as is the case at term in human fetuses. The pancreatic islets of Langerhans were small and not remarkable.
13) Genetics
This species has 38 chromosomes and includes a diminutive Y-chromosome (Jorge et al, 1977, Benirschke, 2006). Most autosomes are metacentric and thus, this appears to be ‘the most derived’ karyotype of the Dasypodidae.
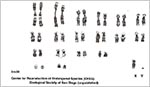 |
Karyotype of three-banded armadillo. |
14) Immunology
No information exists.
15) Pathological features
The soft skin of the neonate’s belly appeared to be green. The lungs of this 2 day-old neonate were congested and had diffuse hemorrhages. In addition, fresh blood was found in the larynx. Cetica et al. (2005) described the presence of several oocytes within the same follicle.
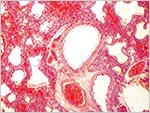 |
Partially inflated lung with fresh hemorrhage. |
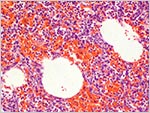 |
Higher power to show the fresh pulmonary hemorrhage with interspersed air bubble. |
16) Physiological features
The neonatal prostate was well developed and is seen next.
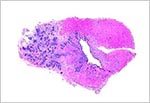 |
Prostate of neonatal three-banded armadillo. |
17) Other resources
This specimen with 38 chromosomes has provided cell strains to the “Frozen Zoo” at CRES, and those of several other three-banded armadillos are available at CRES of the San Diego Zoo. They can be made available to qualified investigators upon request.
18) Other aspects of interest and other needs for study
It would be highly desirable to sample an implanted placenta, discern the depth of trophoblast infiltration, the presence of decidua and the vascular anatomy of placentation.
References
Benirschke, K. p. 81 + in: O’Brien, S.J., Menninger, J.C. and Nash, W.G., eds.: Atlas of Mammalian Chromosomes. Wiley-Liss; A. John Wiley & Sons, Inc. Hoboken, N.J., 2006.
Brinck-Johnsen, T., Benirschke, K. and Brinck-Johnsen, K.: Hormonal steroids in the armadillo, Dasypus novemcinctus. Acta Endocrinol. 56:675-690, 1967.
Brinck-Johnsen, T.: Hormonal steroids in the armadillo, .: Hormonal steroids in the armadillo, Dasypus novemcinctus 2 Oestrone and 17β-oestradiol in pregnancy and their in vitro formation by preparations of placentae, early and late in development. Acta Endocrinol. 63:696-704, 1970.
Cetica, P.D., Marcos, H.J.A. and Merani, M.S.: Morphology of female genital tracts in Dasypodidae (Xenarthra, Mammalia): a comparative survey. Zoomorphology 124:57-65, 2005.
Eisentraut, M.: Biologische Studien im bolivianischen Chaco. III Beitrag zur Biologie der Säugetierfauna. Z. Säugetierk. 8:47-69, 1933.
Gotch, A.F.: Mammals – Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Hayssen, V., van Tienhoven, A. and van Tienhoven, A.: Asdell’s Patterns of Mammalian Reproduction: a Compendium of Species-specific Data. Comstock/Cornell University Press, Ithaca, 1993.
Jorge, W., Meritt, D.A. and Benirschke, K.: Chromosome studies in Edentata. Cytobios 18:157-172, 1977.
Meritt, D.A. Jr.: The development of the La Plata three-banded armadillo Tolypeutes matacus at Lincoln Park Zoo. Int. Zoo Ybk. 11:195-106, 1971.
Meritt, D.A. Jr.: The La Plata three-banded armadillo Tolypeutes matacus in captivity. Int. Zoo Ybk. 16:153-156, 1976.
Puschmann, W.: Zootierhaltung. Tiere in menschlicher Obhut. Säugetiere. Verlag Harri Deutsch, Frankfurt, 2004.
Thenius, E.: Stammesgeschichte der Säugetiere (einschliesslich Hominiden). Handbuch der Zoologie. De Gruyter & Co. Berlin. Vol. 8 (47th Lieferung) 2(1) 1-368, 1969.
Thenius, E. and Hofer, H.: Stammesgeschichte der Säugetiere. Springer-Verlag, Berlin, 1960.
Vizcaíno, S.F. and Loughry, W.J.: The Biology of the Xenarthra. University Press of Florida, Gainesville 2008. (While this is a superb review of many Xenarthran species, Tolypeutes is not covered).
Wetzel, R.M., Gardner, A.L., Redford, K.H. and Eisenberg, J.F.: Order Cingulata, pp. 128-156 in: Gardner, A.L.: Mammals of South America. Vol.1. University of Chicago Press, 2007.
Wilson, D.E. and Reeder, D.A.M.: Mammal Species of the World. A Taxonomic and Geographic Reference. 2nd ed. Smithsonian Institution Press, Washington, DC, 1992.