 |
|
(Clicking
on the thumbnail images below will launch a new window and a larger
version of the thumbnail.)
|
Hemitragus hylocrius
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
There are three recognized species of tahrs: Hemitragus jemlahicus, the Himalayan tahr which is also the best-known species, H. jayakuri, the Arabian or Oman tahr, and H. hylocrius, the Nilgiri tahr from southern India (Nowak, 1999). The animal whose placenta was studied resided at San Diego Zoo, where a successful colony is kept. The term "tahr" is reported to be Nepalese for this species (Gotch, 1979).
Tahrs may have derived from the ancestral form Myotragus balearicus, according to Lalueza-Fox et al. (2000) who studied the mtDNA of that species. Tahrs are said to weight between 50 and 100 kg. The Himalayan tahr has been introduced to New Zealand where it is doing extremely well. Longevity of the Himalayan tahr is over 21 years according to Jones (1993).
 |
Nilgiri Tahr at the San Diego Zoo. |
2) General Gestational Data
The length of gestation is not known precisely. Nowak (1999) gives 177-242 days as a range, much too wide for being the normal length of gestation. Better records need be sought. There is normally one young offspring, rarely two.
3)
Implantation
No implanted placenta has been observed and such a specimen needs also
to be sought in the future. Thus, because no early implantational stages
are known, no decision can be made on the manner of implantation. Perhaps
it is similar to other caprines.
4)
General Characterization of the Placenta
This placenta was significantly autolyzed. It is a polycotyledonary placenta
with 40 cotyledons measuring from 0.8 to 3 cm in diameters. The entire
placenta measured 44 x 23 cm and weighed 110 g.
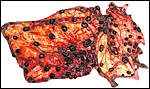 |
Second Nilgiri tahr placenta with 85 cotyledons. |
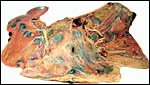 |
Fetal surface of Nilgiri tahr placenta. |
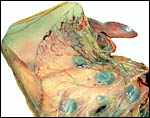 |
The allantoic sac of this placenta had a pronounced vasculature. |
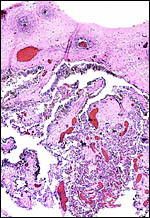 |
Surface of this autolyzed placenta with chorion and ramification of villi. |
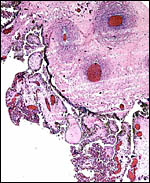 |
Another section of the placental surface with pigmented trophoblast. |
It is likely that this is an epithelial-chorial relation but the placenta was too autolyzed to be definitive about this. The remarkable feature of this organ was the accumulation of pigment in the trophoblast beneath the chorionic plate. This is shown in the next photograph. As in other ungulates, the pigment is accumulated in large chunks within the trophoblast and is also present outside of these cells. It is, again, iron-stain negative. Once again also, this brings up the question of the nature of this pigment. Much more discussion on this matter is written in the chapter on the Alpine Ibex which should be consulted. The pigment is also present beneath the chorion in the Canadian beaver placenta (see that chapter) but it is also iron-stain-negative, while the principal student of that placenta identified iron staining in the inverted yolk sac epithelium of the beaver (Fischer, 1971, 1985). In further searching the literature, the paper by Ishizaki & Belter (1960) was reviewed. These authors identified melanin in human placentas, especially in women with melanosis and skin disorders. Moreover, the rather substantial amounts of melanin they depicted in what they termed "dermatopathic melanosis of the placenta" were also predominantly located in trophoblast, with other sites being the Hofbauer cells (macrophages of villi). The possibility might thus be entertained, in order to explain this pigment in ungulates, that skin disorders in pigmented animals release melanin that thus reaches the placenta. In that event, melanemia might be looked for in future.
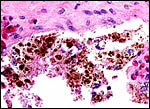 |
As many other ungulate placentas, this specimen had pronounced pigmentation beneath the chorionic surface within and outside the trophoblast. |
The umbilical cord had only 2 blood vessels, but it was badly torn and was probably incompletely represented. All other artiodactyla so far studied have had four vessels and an allantoic duct. The cord was not significantly spiraled, and was 8 cm long and 1.5 cm wide.
7) Uteroplacental circulation
This has not been studied.
8) Extraplacental membranes
Numerous soft, yellowish hippomanes were present. The allantoic membrane had a pronounced vasculature, as shown in the second macroscopic picture but the epithelium was severely autolyzed.
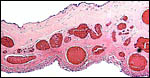 |
Section through the membrane that divides the allantois (below) and amnion above. The epithelium is severely autolyzed, but the extent of allantoic vascularity is apparent. |
No trophoblast external to the endometrium is known to be present and it is not likely to be present, arguing from other related species.
10) Endometrium
This cannot be decided in the absence of an implanted placenta or a pregnant uterus.
11) Various features
There is no subplacenta and no other unusual features are known to me.
12) Endocrinology
I am not aware of any endocrine studies.
13) Genetics
The Nilgiri tahr has 58 chromosomes, as does the Arabian tahr. The Himalayan tahr, however, has only 48 chromosomes (Benirschke & Kumamoto, 1980; see also Nelson-Rees et al., 1967 and Bunch & Nadler, 1980). The latter authors studied Himalayan tahrs with Giemsa banding methods and compared the results with many other caprine species. They concluded from this investigation that chromosomal fusion, rather than fission, best explains the evolutionary relationships and chromosomal rearrangements. Hybrids among tahr species have not been reported to my knowledge. Hybrids of tahrs with goats have resulted in abortions, but in no live offspring (Gray, 1972).
 |
Karyotype of male Nilgiri tahr from the San Diego Zoo. (2n=58). |
Some immunological studies have been published concerning the response to viral infections (see next section).
15) Pathological features
Nielsen et al. (1988) described the deaths of 15 animals due to two outbreaks of presumed malignant catarrhal fever. It caused a severe respiratory illness that led to death in five days. The virus for these outbreaks was not identified, however. There is only a very sparse literature on Nilgiri and Arabian tahr; most reports concern the Himalayan tahr, such as Griner's report (1983) of the occurrence of cleft palate in a newborn. Gazit et al. (1996) reported the occurrence of viral caprine arthritis/encephalitis infection in these animals. They and Yaniv et al. (1985) studied the DNA of this virus. Dahlberg et al. (1981) reported on this disease and on ovine pneumonia in this species.
16) Physiologic data
I know of no publications on Nilgiri tahr physiology.
17) Other resources
The San Diego Zoo's research laboratory, CRES, has many frozen cell strains of Nilgiri tahr, as well as of the other two species, that can be made available by contacting Dr. Oliver Ryder at oryder@ucsd.edu.
18) Other remarks - What additional Information is needed?
Much too little is known of the reproductive physiology of tahrs, including early placentation. A pregnant uterus needs to be studied, endocrinology and physiology need attention.
Acknowledgement
The animal photograph in this chapter comes from the Zoological Society of San Diego. I appreciate also very much the help of the pathologists at the San Diego Zoo.
References
Benirschke, K. and Kumamoto, A.T. The chromosomes of the Nilgiri tahr Hemitragus hylocrius. Intern. Zoo. Yearbk. 20:274-275, 1980.
Bunch, T.D. and Nadler, C.F.: Giemsa-band patterns of the tahr and chromosomal evolution of the tribe Caprini. J. Hered. 71:110-116, 1980.
Dahlberg, J.E., Gaskin, J.M. and Perk, K.: Morphological and immunological comparison of caprine arthritis encephalitis and ovine progressive pneumonia viruses. J. Virol. 39:914-919, 1981.
Fischer, T.V.: Placentation in the American beaver (Castor canadensis). Amer. J. Anat. 131:159-184, 1971.
Fischer, T.V.: The subplacenta of the beaver (Castor canadensis). Placenta 6:311-321, 1985.
Gazit, A., Mashiah, P., Kalinski, H., Gast, A., Rosin-Abersfeld, R. Tronick, S.R. and Yaniv, A.: Two species of Rev proteins, with distinct N termini, are expressed by caprine arthritis encephalitis virus. J. Virol. 70:2674-2677, 1996.
Gotch, A.F.: Mammals - Their Latin Names Explained. Blandford Press, Poole, Dorset, 1979.
Gray, A.P.: Mammalian Hybrids. A Check-list with Bibliography. 2nd edition. Commonwealth Agricultural Bureaux Farnham Royal, Slough, England, 1972.
Griner, L.A.: Pathology of Zoo Animals. Zoological Society of San Diego, San Diego, California, 1983.
Ishizaki, Y. and Belter, L.F.: Melanin deposition in the placenta as a result of skin lesions (dermatopathic melanosis of placenta). Amer. J. Obstet. Gynecol. 79:1074-1077, 1960.
Jones, M.L.: Longevity of ungulates in captivity. Intern. Zoo Yearbk. 32:159-169, 1993.
Lalueza-Fox, C., Bertranpetit, J., Alcover, J.A., Shailer, N. and Hagelberg, E.: Mitochondrial DNA from Myotragus balearicus, an extinct bovid from the Balearic Islands. J. Exp. Zool. 288:56-62, 2000.
Nelson-Rees, W.A., Kniazeff, A.J., Malley, R.L. and Darby, N.B. Jr.: On the karyotype of the tahr Hemitragus jemlahicus and the Y-chromosome of goats and sheep. Chromosoma 23:154-161, 1967.
Nielsen, N.O., Oosterhuis, J., Janssen, D., McColl, K., Anderson, M.P. and Heuschele, W.P.: Fatal respiratory disease in Nilgiri tahr: possibly malignant catarrhal fever. Can. J. Vet. Res. 52:216-221, 1988.
Nowak, R.M.: Walker's Mammals of the World. 6th ed. The Johns Hopkins Press, Baltimore, 1999.
Yaniv, A., Dahlberg, J.E., Tronick, S.R., Chiu, I.M. and Aaronson, S.A.: Molecular cloning of integrated caprine arthritis-encephalitis virus. Virology 145:340-345, 1985.